به ساخت یک مولکول RNA با الگوگیری از توالی DNA، رونویسی میگویند. فرآیند رونویسی به طور کلی شامل سه مرحله، آغاز رونویسی، طویلسازی و خاتمه رونویسی است. عوامل متعددی در این فرآیند وظایف منحصر به فرد خود را دارند. در این مطلب از مجله فرادرس به سراغ مراحل رونویسی و عوامل موثر در هر مرحله میرویم، با آنزیم RNA پلیمراز، فاکتورهای رونویسی و توالیهای تنظیمی مانند افزایندهها و خاموشگرها آشنا میشویم. در نهایت نیز با استفاده از یک جدول شباهتها و تفاوتهای رونویسی در پروکاریوتها و یوکاریوتها را بررسی میکنیم.


مراحل رونویسی
در زیستشناسی به فرآیند تهیه رونویسی از اطلاعات ذخیره شده در رشته DNA که باعث تولید یک رشته RNA مکمل با DNA میشود، «فرآیند رونویسی» میگوییم. در پروکاریوتها فرآیند رونویسی در سیتوپلاسم و در یوکاریوتها درون هسته سلول انجام میشود. به طور کل رونویسی شامل سه مرحله است.
- آغاز
- طویلسازی
- خاتمه
رونویسی اولین مرحله از مراحل «بیان ژن» (Gene Expression) است که در طی آن با استفاده از اطلاعات ژنتیکی سلول، محصولاتی دارای فعالیت، مانند پروتئینها ساخته میشوند. با توجه به تعریفی که از رونویسی ارائه دادیم، میتوان گفت که هدف از رونویسی ساخت یک کپی از توالی DNA است که به دلایل مختلف ازجمله استفاده از نوکلئوتیدهای متفاوت به صورت RNA ساخته میشود. در صورتی که تمایل به کسب اطلاعات کاملتر راجع به ساختار و انواع مولکولهای RNA دارید، مطالعه مطلب «RNA چیست؟ — به زبان ساده» از مجله فرادرس را به شما پیشنهاد میکنیم.
RNA پلیمراز با تعدادی از فاکتورهای رونویسی به بخش پروموتور در DNA متصل میشود. این اتصال باعث باز شدن پیچ DNA و شکلگیری حباب رونویسی میشود.

مرحله آغاز رونویسی
RNA پلیمراز با کمک فاکتورهای رونویسی خاصی به پروموتور متصل میشود و «کمپلکس بسته RNA پلیمراز-پروموتور» (RNA Polymerase-Promoter Closed Complex) را میسازد. در ناحیه این کمپلکس، دو رشته DNA به هم متصل هستند و هنوز پیوندهای هیدروژنی بین آنها باز نشده است. توالیهای پروموتور در ابتدای ژنها قرار دارند اما در یوکاریوتها و پروکاریوتها در مورد توالی پروموتور یک تفاوت وجود دارد.
- یوکاریوتها: هر ژن پروموتور اختصاصی خود را دارد.
- پروکاریوتها: ممکن است چند ژن که با یکدیگر رونویسی میشوند، یک پروموتور داشته باشند.
به محض اتصال RNA پلیمراز، این آنزیم باعث جدا شدن دو رشته DNA از یکدیگر میشود و به این ترتیب تکرشته الگو در دسترس این آنزیم قرار میگیرد. اگر برای شما سوال است که منظور از رشته الگو چیست؟ باید بگوییم که یکی از رشتههای DNA به عنوان الگویی برای RNA پلی مراز عمل میکند تا بتواند مولکول RNA را بسازد، به این رشته «رشته الگو» (Template Strand) یا «رشته غیر کدکننده» (Noncoding Strand) میگوییم. به ناحیه حضور RNA پلیمراز نیز که دو رشته DNA در آن ناحیه باز شدهاند «حباب رونویسی» (Transcription Bubble) میگویند.

پس از تشکیل حباب رونویسی، RNA پلیمراز به همراه یک یا چند فاکتور رونویسی جایگاه شروع رونویسی را پیدا میکند. با یافتن این جایگاه اولین نوکلئوتید را گذاشته و سپس شروع به ساخت محصول RNA اولیه میکند. مرحله آغاز رونویسی توسط پروتئینهای دیگری نیز تنظیم میشود که به آنها «فعالکننده» (Activator) و «سرکوبگر» (Repressor) میگوییم. این پروتئینها قادر به تنظیم و متعادلسازی تشکیل و فعالیت کمپلکس آغاز رونویسی هستند.
یکی از نکات بسیار مهم که تفاوت رونویسی و همانندسازی در مرحله آغازین را نشان میدهد، عدم نیاز RNA پلیمراز به پرایمر است در حالی که در فرآیند همانندسازی DNA، DNA پلیمراز نیاز به پرایمر دارد. در صورتی که تمایل دارید تا اطلاعات بیشتر و بهتری در مورد همانندسازی به دست آورید، مطالعه مطلب «همانند سازی DNA — به زبان ساده» از مجله فرادرس را به شما توصیه میکنیم.
پروموتور چیست؟
محل آغاز همانندسازی توسط ناحیهای به نام «پروموتور» (Promoter) مشخص میشود. پروموتور محل اتصال RNA پلیمراز است و با اتصال آنزیم به این ناحیه، رونویسی شروع میشود؛ بنابراین حضور پروموتور برای شروع مراحل رونویسی ضروری است. پروموتورهای یوکاریوتی و پروکاریوتی تفاوتهایی باهم دارند که در این بخش از بررسی مراحل رونویسی به معرفی هر دو آنها میپردازیم.
پروموتورهای پروکاریوتی
پروموتورهای پروکاریوتی در ناحیه ۳۵- و ۱۰- قرار دارند و توسط فاکتور سیگمای RNA پلیمراز تشخیص داده میشوند. آنزیم RNA پلیمراز با اتصال به پروموتور «کمپلکس بسته پروموتور» (Closed Promoter Complex) را شکل میدهد. دو ناحیه ۳۵- و ۱۰- توالیهای نوکلئوتیدی منحصر به فرد خود را دارند که توسط آنزیم شناسایی میشود، در ادامه توالی مربوط به هر ناحیه را آوردهایم.
- توالی ناحیه ۳۵- پروموتور پروکاریوتی به ترتیب روی رشته الگو و رشته کدکننده: AACTGT و TTGACA
- توالی ناحیه ۱۰- پروموتور پروکاریوتی به ترتیب روی رشته الگو و رشته کدکننده: ATATTA و TATAAT

پروموتورهای یوکاریوتی
در یوکاریوتها برخلاف پروکاریوتها، RNA پلیمراز به طور مستقیم به توالی پروموتور متصل نمیشود. در ابتدا فاکتوری پروتئینی به نام «فاکتور عمومی رونویسی» (General Transcription Factor | GTF) به توالی پروموتور متصل میشود و به این ترتیب به RNA پلیمراز برای اتصال به رشته DNA الگو کمک میکند.
یوکاریوتها توالی پروموتوری به نام «TATA Box» دارند که توسط فاکتورهای رونویسی شناسایی میشود و به RNA پلیمراز اجازه اتصال به DNA را میدهد. در ناحیه TATA Box نوکلئوتیدهای آدنین و تیمین حضور پر رنگی دارند که باعث میشود دو رشته DNA به سادگی از یکدیگر جدا شوند. اگر اولین نوکلئوتیدی که رونویسی میشود را ۱+ در نظر بگیریم، TATA Box حدود ۲۵ نوکلئوتید قبل از اولین نوکلئوتید قرار دارد و موقعیت آن را به صورت ۲۵- نشان میدهیم.

عبور از پروموتور
پس از تشکیل اولین پیوند، RNA پلیمراز باید از توالی پروموتور عبور کند. در این زمان RNA پلیمراز تمایل به رها کردن رشته کوتاه RNA ساخته شده دارد، این تمایل تا زمانی که رشته RNA به طول حدود ۱۰ نوکلئوتید برسد وجود خواهد داشت.
مسیر یادگیری ژنتیک مولکولی با فرادرس
در سالهای اخیر ژنتیک مولکولی تبدیل به یکی از مهمترین بخشهای زیستشناسی شده است و محققان فعال در شاخههای مختلف بخش از تحقیقات خود را در سطح مولکولی انجام داده و گزارش میکنند. به کمک تکنولوژیهای جدید اطلاعات مختلفی راجع به توالی ژنتیکی و پروتئينی جانداران مختلف در دسترس است که میتوانند برای اهداف تحقیقاتی مختلف استفاده شوند.
برای آشنایی با ژنتیک مولکولی ابتدا باید با مفاهیم پایهای ژنتیک آشنا شد و سپس به سراغ فرآیندهای مختلف در سطح ژنوم موجودات رفت، پس از کسب این اطلاعات میتوان قدمی فراتر گذاشت و با پردازش اطلاعات به دست آمده به سراغ طراحی پروتئینهای نوترکیب، شناسایی ژنهای مسئول در بیماریهایی مختلف و غیره رفت. فرادرس دورههای متفاوتی طراحی، تهیه و منتشر کرده است که نیازهای موجود در این مسیر را پوشش میدهند، در ادامه تعدادی از این دورهها را به شما معرفی میکنیم.

فرادرس با دنبال کردن علم روز دنیا در دنیای بیوتکنولوژی و دادههای زیستی، دورههای متنوع و کاربردی در زمینه الگوریتمهای ژنتیکی و محاسبات تکاملی تهیه و منتشر کرده است که برای دیدن تمام دورهها بازدید از صفحه مجموعه فیلمهای آموزش الگوریتم ژنتیک و محاسبات تکاملی – مقدماتی تا پیشرفته فرادرس را به شما پیشنهاد میکنیم.
مرحله طویلسازی رونویسی
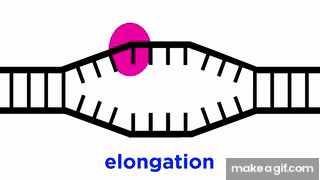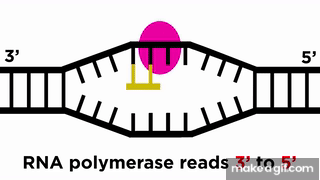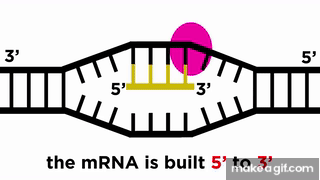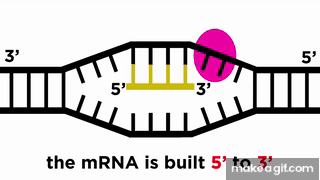
RNA پلیمراز نوکلئوتید به نوکلئوتید روی رشته الگو جلو میرود و با خواندن توالی رشته الگو، RNA را میسازد. حرکت RNA پلیمراز روی رشته الگو مانند حرکت یک قطار روی ریل است، پس میتوان انتظار داشت که جهت حرکت قطار نیز مهم باشد. رشته RNA در جهت به () ساخته میشود، یعنی نوکلئوتید جدید از ناحیه فسفات خود به نوکلئوتید از قبل موجود در انتهای رشته متصل میشود.
توالی رشته RNA مشابه رشته مکمل رشته الگوی است که به آن «رشته کدکننده» (Coding Strand) نیز میگویند. البته باید این نکته را در نظر داشت که در RNA به جای نوکلئوتید تیمین (T) از یوراسیل (U) استفاده میشود.

برای ساخته شدن مولکول RNA در جهت ، باید RNA پلیمراز در جهت رشته DNA الگو حرکت کند. در کادر موجود در تصویر بالا، میتوان هر دو این جهتها را شناسایی کرد. همانطور که در این تصویر میبینیم، جهتگیری مولکول RNA با رشته کدکننده DNA مشابه است، پیشتر هم گفتیم که هر دو این رشتهها توالی یکسانی دارند، اما این دو رشته ۲ تفاوت هم دارند که در ادامه به آنها اشاره میکنیم.
- مولکول قند به کار رفته در ساختار RNA، قند پنج کربنه ریبوز و در DNA دئوکسیریبوز است.
- در DNA نوکلئوتید تیمین وجود دارد، در حالی که در RNA نوکلئوتید یوراسیل حضور دارد.
در هنگام رونویسی، امکان دارد که تعداد زیادی RNA پلیمراز به رشته DNA الگو متصل شوند و شروع به رونویسی و تولید mRNA کنند، بنابراین به کمک این روش از روی یک نسخه ژن در مدت بسیار کوتاهی، تعداد زیادی mRNA تولید میشود.
سرعت مرحله طویلسازی در پروکاریوتها و یوکاریوتها در حدود ۱۰ الی ۱۰۰ نوکلئوتید بر ثانیه است. البته باید در نظر داشت که در یوکاریوتها، نوکلئوزومها مانند سدهایی بر سر راه RNA پلیمرازها قرار دارند و میتوانند روی سرعت حرکت و فعالیت آن ها تاثیر بگذارند. در این جانداران، افت سرعتی که به خاطر نوکلئوزومها رخ میدهد، توسط «فاکتورهای طویلسازی» (Transcription Elongation Factors) مانند TFIIS تنظیم میشود.
مرحله طویلسازی شامل مکانیسمهای بازبینی برای اصلاح نوکلئوتیدهایی است که به اشتباه در توالی RNA قرار میگیرند. در یوکاریوتها این روند ممکن است باعث وقفهای کوتاه در رونویسی شود، زیرا باید فاکتورهای مربوط به اصلاح RNA به سیستم اضافه شوند. این وقفه ممکن است به دلیل RNA پلیمرازها یا به خاطر ساختار کروماتین باشد.
منظور از چیست؟
رشتههای RNA و DNA دارای دو انتها هستند که با یکدیگر تفاوت دارند. این تفاوت باعث سوگیری یا جهتدهی DNA و RNA میشود. در ادامه خصوصیت هر انتها را توضیح خواهیم داد تا تفاوت آنها را بهتر متوجه شویم.
- «انتهای » ( end): در این انتها، گروه فسفات نوکلئوتیدی که سر رشته قرار دارد، برای برقراری پیوند فسفو دیاستری استفاده نشده و آزاد است. این گروه فسفات به پنجمین کربن موجود در حلقه قند پنج کربنه متصل است، به همین دلیل است که به این انتها، «سر » میگوییم.
- «انتهای » (end): در انتهای یک رشته، گروه هیدروکسیل آخرین نوکلئوتید موجود در رشته، آزاد است. گروه هیدروکسیل به سومین کربن موجود در حلقه قند متصل است و به همین دلیل به این انتها «سر » میگوییم.
یکی از دلایل اهمیت آشنایی با جهت حرکت آنزیمها و جهتگیری رشتهها این است که بسیاری از فرآیندها، مانند رونویسی و همانندسازی DNA، تنها در یک جهت انجام میشوند.

مرحله خاتمه رونویسی
تا اینجا با دو مرحله از مراحل رونویسی آشنا شدیم، در این بخش به مرحله آخر رونویسی میپردازیم. در پروکاریوتها در انتهای ناحیه رونویسی، توالیهایی وجود دارند که پیام خاتمه رونویسی را منتقل میکنند، به این توالیها «پایانگر» (Terminator) میگویند. زمانی که RNA پلیمراز این توالیها را رونویسی کند، محصول رونویسی (RNA) از RNA پلیمراز رها میشود. یکی از مثالهای مکانیسمهای خاتمه رونویسی، تشکیل ساختارهای «سنجاق سری» (Hairpin) در RNA است که در تصویر زیر نیز میتوانید تشکیل آن را ببینید.

باکتریها از دو استراتژی متفاوت برای پایان رونویسی استفاده میکنند.
- پایان غیروابسته به Rho: رونویسی RNA زمانی متوقف میشود که مولکول RNA تازه سنتز شده، یک ساختار سنجاق سری غنی از C-G (نوکلئوتیدهای سیتوزین و گوانین) تشکیل دهد که پس از آن در رشته RNA تعداد زیادی یوراسیل وجود دارد. زمانی که ساختار سنجاقسری شکل میگیرد، فشار وارد شده توسط این ساختار پیوندهای ضعیفی که بین یوراسیلهای رشته RNA و آدنینهای رشته DNA وجود دارند را از بین میبرد و به این ترتیب RNA از DNA جدا میشود و رونویسی به پایان میرسد.
- پایان وابسته به Rho: پروتئین Rho فاکتوری با قابلیت ناپایدار کردن ارتباط بین الگو و mRNA است، بنابراین باعث آزاد شدن mRNA تازه سنتز شده میشود.
مکانیسمهای پایان رونویسی در یوکاریوتها نسبت به باکتریها کمتر شناخته شدهاند، اما شامل برش در RNA جدید و افزودن دمی از نوکلئوتیدهای آدنین به انتهای است که به آن «دم پلی A» میگوییم. این فرآیند «پلی آدنیلاسیون» (Polyadenylation) نام دارد.

روش پایان رونویسی در یوکاریوتها با پروکاریوتها متفاوت است. در حقیقت در یوکاریوتها توالی وجود ندارد که به RNA پلیمراز ۲ نشان دهد که رونویسی باید متوقف شود. در نتیجه ممکن است RNA پلیمراز ۲ رونویسی را تا صدها یا هزاران نوکلئوتید پس از نقطه پایان ادامه دهد.
رونوشت ساخته شده قبل از پایان رونویسی RNA پلیمراز ۲ توسط آنزیمهای خاصی برش میخورد و آزاد میشود تا به عنوان mRNA نابالغ وارد مراحل پردازش پیش از ترجمه شود. ناحیه برش به عنوان پایان ژن در نظر گرفته میشود و بخشهای اضافی که توسط RNA پلیمراز ساخته شدند توسط «-اگزونوکلئاز» تجزیه میشوند. این آنزیم با تجزیه RNA اضافی که توسط RNA پلیمراز ساخته میشود، خود را به آنزیم RNA پلیمراز میرساند و باعث جدا شدن آن از DNA میشود.

تغییرات پیش از ترجمه mRNA
پس از طی کردن مراحل رونویسی از RNA، DNA تولید میشود که در پروکاریوتها بدون هیچ تغییری برای ترجمه استفاده میشود اما در یوکاریوتها به mRNA تولید شده در فرآیند رونویسی «mRNA نابالغ» یا «mRNA اولیه» (pre-mRNA) گفته میشود. این mRNA باید فرآیندی را تحت عنوان «بلوغ mRNA» طی کند و تبدیل به «mRNA بالغ» (Mature mRNA) شود. فرآیند بالغ شدن mRNA سه مرحله دارد که در ادامه از آنها را توضیح میدهیم.
- کلاهکگذاری سر : در این مرحله یک کلاهک گوانین متیله شده به سر mRNA اضافه میشود. کلاهک سر به تشخیص مولکول mRNA توسط ریبوزوم کمک میکند، همچنین از mRNA نابالغ در برابر تخریب حاصل از حمله RNAseها که آنزیمهای تجزیهکننده RNA هستند، محافظت میکند.
- پلی آدنیلاسیون: در طی مرحله یک دم پلی A به سر RNA اضافه میشود. دم پلی A از تعداد زیادی مولکول «آدنوزین مونوفسفات» ساخته شده است. این دم باعث تثبیت مولکول RNA میشود.
- پیرایش RNA: پیرایش RNA در حین رونویسی اتفاق میافتد و باعث میشود که از یک mRNA نابالغ انواع مختلفی پروتئین ساخته شود. پیرایش RNA ۲ مرحله دارد که در ادامه آنها را توضیح دادیم.
- حذف بخشهای غیرکدکننده که با عنوان «اینترون» (Intron) شناخته میشوند.
- اتصال بخشهای حاوی کدژنتیکی که با عنوان «اگزون» (Exon) شناخته میشوند.

عوامل رونویسی
عوامل متعددی در فرآیند پیچیده رونویسی فعالیت دارند. اصلیترین آنزیم رونویسی، RNA پلیمراز است که در پروکاریوتها یک نوع و در یوکاریوتها سه نوع متفاوت از آن وجود دارد. علاوه بر این آنزیم مهم که زیرواحدهای گوناگونی دارد، فاکتورهای مختلفی روی فرآیند رونویسی تاثیرگذارند. برای مثال بعضی از این فاکتورها به اتصال آنزیم RNA پلیمراز به پروموتورهای ژنها کمک میکنند و بعضی دیگر ممکن است با جلوگیری از این اتصال، مانع ساخت RNA شوند. در این بخش با این عوامل آشنا میشویم تا درک بهتری از فعالیتهای آنها در مراحل رونویسی داشته باشیم.
RNA پلیمراز
آنزیم اصلی فرآیند رونویسی «RNA پلیمراز» (RNA Polymerase) است که از DNA تکرشتهای به عنوان الگو استفاده میکند تا RNA مکمل آن را بسازد. RNA پلیمراز به طور خاص RNA را در جهت به میسازد. منظور از این جهت این است که هر نوکلئوتید جدیدی که به رشته در حال ساخت اضافه میشود به قسمت نوکلئوتید قبلی متصل میشود. آنزیم RNA پلیمراز، فاکتوری به نام «فاکتور سیگما» دارد که بخشی از آنزیم است که قابلیت شناسایی توالی پروموتور را دارد.

یوکاریوتها سه نوع RNA پلیمراز دارند، در حالی که پروکاریوتها کل فرآیند رونویسی را تنها با یک آنزیم RNA پلیمراز پیش میبرند. هر نوع از RNA پلیمرازهای یوکاریوتی مسئول سنتز RNAهای خاصی هستند که در ادامه با آنها آشنا میشویم.
- RNA پلیمراز ۱: در هسته قرار دارد و زیرواحدهای ۲۸S، ۱۸S و ۵٫۸S متعلق به rRNA (RNA ریبوزومی) را میسازد.
- RNA پلیمراز ۲: در هسته قرار دارد و از روی ژنهای کدکننده پروتئینها رونویسی میکند. محصول فعالیت این RNA، mRNA است. ژنهای کدکننده RNAهای کوچک (مانند snoRNAs و snRNAs به جز U6 snRNA) توسط این RNA پلیمراز رونویسی میشوند.
- RNA پلیمراز ۳: این نوع RNA پلیمراز که مانند دو مورد قبلی در هسته قرار دارد، ژنهای مربوط به tRNA، U6 snRNA، ۵S rRNA و ۷S RNA را رونویسی میکند.
هر سه این آنزیمها از حدود ۱۲ زیرواحد ساخته شدهاند که این موضوع نشان میدهد این RNA پلیمرازها کمپلکسهای بسیار بزرگ آنزیمی هستند. با توجه به توضیحات بالا میتون گفت که در ساخت آنزیمهای پروتئينی، RNA پلیمراز ۲ فعالیت دارد و بخش اعظمی از ریبوزومها نیز توسط RNA پلیمراز ۱ ساخته میشود.
فاکتورهای رونویسی
در زیستشناسی مولکولی، «فاکتور رونویسی» (Transcription Factor | TF) پروتئینی است که سرعت رونویسی DNA و ساخت «RNA پیامرسان» (Messenger RNA) را با اتصال به توالیهای خاصی از DNA، کنترل میکند. وظیفه فاکتور رونویسی روشن کردن یا خاموش کردن ژنها است.
بسیاری از ژنها فقط در سلولهای خاصی بیان میشوند. این که چه ژنی در چه نوع سلولی بیان شود، توسط فاکتورهای رونویسی کنترل میشود. زمان و میزان بیان این ژنها نیز مهم است، این موضوع نیز توسط فاکتورهای رونویسی در طول زندگی سلول تنظیم میشود.
تعداد فاکتورهای رونویسی پیدا شده در یک جاندار با توجه به اندازه ژنوم موجود تغییر میکند، به این صورت که با افزایش اندازه ژنوم، تعداد فاکتورهای رونویسی نیز افزایش مییابند. حدود ۲۸۰۰ پروتئين در انسان شناسایی شدند که «دومین» (Domain) اتصال به DNA را دارند و به نظر میرسد حدود هزار و هشتصد پروتئین از آنها به عنوان فاکتور رونویسی فعالیت میکنند. بنابراین میتوان نتیجه گرفت که حدود ۱۰ درصد ژنوم، انواع مختلف فاکتورهای رونویسی را کد میکند.

مکانیسم فعالیت فاکتورهای رونویسی
فاکتورهای رونویسی به نواحی «پروموتور» (Promoter) یا «افزایشدهنده» (Enhancer) که روی DNA قرار دارند، متصل شده و بیان ژنها را تنظیم میکنند. با توجه به نوع فاکتور رونویسی، میزان رونویسی از ژن مدنظر افزایش یا کاهش مییابد. فاکتورهای رونویسی از روشهای متفاوتی برای تنظیم بیان ژن استفاده میکنند که در ادامه از تعدادی از آنها نام میبریم.
- تثبیت یا جلوگیری از اتصال RNA پلیمراز به DNA.
- استیلاسیون یا استیلزدایی از پروتئينهای هیستون. فاکتورهای رونویسی میتوانند این کار را به تنهایی انجام دهند یا از پروتئينهای دیگر برای پیشبرد این فعالیت استفاده کنند.
- استیله کردن هیستونها: استیله کردن هیستونها باعث میشود که ارتباط این پروتئینها با رشته DNA ضعیف شود، بنابراین DNA برای رونویسی در دسترستر خواهد بود.
- استیلزدایی از هیستونها: استیلزدایی از هیستونها، ارتباط DNA با هیستونها را تقویت میکند و به این ترتیب DNA برای رونویسی در دسترس RNA پلیمراز نیست.
- استفاده از «کمکفعال کننده» (Coactivator) یا «کمکسرکوبگرها» (Corepressor) برای کمپلکس رونویسی DNA.
به توالی DNA که فاکتورهای رونویسی به آن متصل میشوند «جایگاه اتصال فاکتور رونویسی» میگویند.
توالی تنظیمی چیست؟
تا اینجای این مطلب از مجله فرادرس پس از آشنایی با مراحل رونویسی با تغییرات پیش از ترجمه mRNAهای یوکاریوتی نیز آشنا شدیم، اما در فرآیند رونویسی ژنها، توالیهای دیگری نیز نقش پررنگی دارند که در این بخش به آنها میپردازیم. «توالی تنظیمی» (Regulatory Sequence) به توالی DNA گفته میشود که با تشکیل کمپلکسهای DNA-پروتئين، بیان ژنها را کنترل میکند. پروتئينهایی که در این فرآیند نقش دارند، توانایی اتصال به DNA را دارند. این توالیها توانایی افزایش یا کاهش بیان ژنهای موجودات زنده را دارند. در حقیقت باید گفت که تنظیم بیان ژن یکی از حیاتیترین فرآیندهایی است که در تمام موجودات زنده و حتی ویروسها پیگیری میشود.
پروموتورها، «افزاینده» (Enhancer)، «خاموشگر» (Silencer) و دیگر بخشهایی از DNA که پروتئینهای تنظیمی به آنها متصل میشوند، ازجمله توالیهای تنظیمی هستند. توالیهای تنظیمی را میتوان روی mRNA هم دید اما این دسته از توالیهای تنظیمی به اندازه توالیهایی که روی DNA قرار دارند، هدف مطالعه قرار نگرفتهاند.
در DNA، به طور معمول تنظیم بیان ژن در سطح بیوسنتز RNA (رونویسی) انجام میشود. این فرآیند توسط اتصال پروتئینهایی که در بخش عوامل رونویسی با آنها آشنا شدیم، یعنی فاکتورهای رونویسی، کامل میشود. این پروتئینها توانایی فعال یا مهار کردن رونویسی را دارند. فاکتورهای رونویسی ممکن است به عنوان فعالکننده، مهارکننده یا هر دو عمل کنند.
یک توالی تنظیمی که روی DNA قرار دارد تا زمانی که فعال نشود، توانایی تنظیم کردن روند رونویسی و بیان ژن را ندارد. در اصل توالیهای تنظیمی متفاوتی فعالی میشوند و سپس با استفاده از مکانیسمهای متفاوتی مسیر بیان ژن را تنظیم میکنند.
افزایندهها
افزایندهها توالیهایی از ژنوم هستند که اصلیترین بخش تنظیم بیان ژن به حساب میآیند. افزایندهها برنامه کلی بیان ژن هر نوع از انواع مختلف سلولها را کنترل میکنند. محل قرارگیری افزایندهها ممکن است فاصله بسیار زیادی با ژن هدف داشته باشند (برای مثال ۱ میلیون جفت باز فاصله بین ژن و افزاینده) اما این توالیها با تشکیل یک لوپ خود را به ژن مدنظر میرسانند. محل قرارگیری افزایندهها ممکن است در توالیهای بالادست ژن یا پایین دست باشد. در تصویر زیر میتوانید یک لوپ ایجاد شده توسط افزاینده را ببینید.

طول افزایندهها روی DNA در حدود ۵۰ الی ۱۵۰۰ جفت نوکلئوتید است، این توالیها توانایی اتصال به پروتئینها را دارند؛ فاکتورهای رونویسی همان پروتئينهای تنظیمی هستند که به افزایندهها متصل میشوند. این توالیها هم در سلولهای یوکاریوتی و هم در سلولهای پروکاریوتی وجود دارند و امکان رونویسی از آنها نیز وجود دارد. محصول حاصل از رونویسی این توالیها برای تولید پروتئین استفاده نمیشود و در قالب RNA غیرکدکننده نقش تنظیمی دارد. بیان این نوع از RNAها ا سطح تولید mRNA ژن هدف افزاینده تنظیم میشود.
خاموشگرها
به توالیهایی که پروتئینهای سرکوبگر به آنها متصل میشوند، «خاموشگر» میگویند. پروتئينهای سرکوبگر ازجمله فاکتورهای تنظیم رونویسی هستند. زمانی که این دسته از پروتئينها به خاموشگرها متصل میشوند، جلوی اتصال RNA پلیمراز گرفته میشود و از ژن مربوط به آن خاموشگر رونویسی نمیشود. با غیرفعال شدن رونویسی، پروتئین کد شده توسط آن ژن نیز تولید نمیشود، بنابراین میتوان نتیجه گرفت که خاموشگرها توانایی جلوگیری کردن از سنتز یک پروتئین را دارند.

مهارکنندگان رونویسی
تا اینجای این مطلب از مجله فرادرس با مراحل رونویسی آشنا شدیم، در این بخش قصد داریم با مهارکنندگان این فرآیند مهم سلولی آشنا شویم. مهارکنندگان رونویسی عواملی هستند که برای مهار فعالیت RNA پلیمراز استفاده میشوند و به این ترتیب مانع انجام فرآیند رونویسی میشوند. از مهارکنندگان رونویسی برای متوقفسازی رونویسی در باکتریهای بیماریزا استفاده میشود، در ادامه تعدادی از رایجترین انواع این مهارکنندگان را معرفی میکنیم.
- «آلفا-آمانیتین» (α-Amanitin): مهارکنندهای که از مخمرها استخراج شده است. به طور اختصاصی RNA پلیمراز ۲ و ۳ را مهار میکند.
- «ریفامپین» (Rifampicin): رونویسی در باکتریها را متوقف میکند.
- متلاسیون هیستون: این مکانیسم با تغییر بار هیستون میتواند جلوی فعالیت رونویسی را بگیرد.
جدول مقایسه رونویسی در پروکاریوتها و یوکاریوتها
هنگام آشنایی با مراحل رونویسی از شباهتها و تفاوتهای موجود میان یوکاریوتها و پروکاریوتها گفتیم اما در این بخش قصد داریم به کمک یک جدول بخشهای مختلف رونویسی را در هر دو نوع این سلولها مرور کنیم.
| شباهتها | تفاوتها | |
| DNA به عنوان الگوی رونویسی استفاده میشود. | پروموتور | پروکاریوتها سه بخش مرتبط با پروموتور دارند. (نواحی بالا دست، نواحی ۳۵- و ۱۰-) |
| یوکاریوتها دارای بخشهای متعدد مرتبط با پروموتور هستند. (مانند TATA Box) | ||
| RNA پلیمراز | پروکاریوتها ۱ نوع RNA پلیمراز دارند. | |
| RNA پلیمراز آنزیم اصلی تمام فرآیند رونویسی است. | یوکاریوتها ۳ نوع RNA پلیمراز دارند. | |
| کمپلکس آغازین | یوکاریوتها کمپلکس آغازینی دارند که از فاکتورهای متنوعی ساخته شده است. | |
| پروکاریوتها کمپلکس آغازین ندارند. | ||
| محصول نهایی پروسه رونویسی RNA است. | رونویسی و ترجمه | در پروکاریوتها قبل از اتمام رونویسی، ترجمه شروع میشود. |
| در یوکاریوتها باید رونویسی کامل شود تا ترجمه اتفاق بیفتند. | ||
| ساختار RNA | mRNA پروکاریوتی امکان دارد که حاصل رونویسی از چند ژن باشد. | |
| ساختار شیمیایی رونویسی در هر دو نوع سلول یکسان است. | mRNA یوکاریوتی محصول رونویسی از تنها ۱ ژن است. | |
| خاتمه رونویسی | در پروکاریوتها دو روش وابسته به فاکتور Rho و غیروابسته به فاکتور Rho وجود دارد. | |
| در یوکاریوتها توالی برای القا خاتمه رونویسی وجود ندارد. | ||
جمعبندی
در این مطلب از مجله فرادرس با مراحل رونویسی و عوامل موثر بر آن آشنا شدیم. در جدول زیر خلاصهای از اتفاقات مهم در هر سه مرحله اصلی رونویسی را به طور خلاصه ارائه دادهایم.
| مراحل رونویسی | |
| آغاز رونویسی | RNA پلیمراز و فاکتورهای رونویسی به پروموتور متصل میشوند و پس از تشکیل حباب رونویسی، ساخت RNA شروع میشود. |
| طویل سازی | آنزیم RNA پلیمراز در طول رشته الگو حرکت میکند و RNA را میسازد. |
| خاتمه رونویسی | خاتمه رونویسی در باکتریها به دو مدل «وابسته به فاکتور Rho» و «غیروابسته به فاکتور Rho» تقسیم میشود. |
| در یوکاریوتها توالی خاصی که به RNA پلیمراز خاتمه رونویسی را القا کند وجود ندارد. | |
جهت ساخت mRNA است و جهت حرکت آنزیم RNA پلیمراز از سر رشته الگو به سمت سر است.
رونویسی در یوکاریوتها با سه نوع متفاوت آنزیم RNA پلیمراز انجام میشود. هر آنزیم مسئول ساخت یک نوع RNA است، mRNA را RNA پلیمراز ۲ میسازد. در مقابل پروکاریوتها تنها یک نوع RNA پلیمراز دارند و حتی برای ژنهایی که باهم بیان میشوند یک پروموتور نیز دارند اما در یوکاریوتها هر ژن به تنهایی پروموتور اختصاصی خود را دارد.
در پروکاریوتها همزمان با رونویسی، فرآیند ترجمه نیز آغاز میشود، اما در یوکاریوتها پس از اتمام کار RNA پلیمراز، مولکول RNA نابالغ باید مراحل متفاوتی را طی کند تا بتواند از هسته سلول خارج شده و در سیتوپلاسم ترجمه شود.
source