مولکولهای آلی ترکیباتی هستند که ساختار اصلی آنها از اتمهای کربن تشکیل شده است. این مولکولها در تشکیل بسیاری از ساختارهای زیستی ازجمله غشای سلول و اندامکها، انتقال پیام بین سلولها و تامین انرژی لازم برای فعالیتهای سلول شرکت میکنند. پروتئینها، اسیدنوکلئوئیکها، لیپیدها و کربوهیدرات درشتمولکولهای آلی مهم در ساختارهای زیستی هستند. پروتئینها از زیرواحدهای آمینواسیدی، اسیدنوکلئوئیکها از زیرواحدهای نوکلئوتیدی، کربوهیدراتها از حلقههای کربنی و لیپیدها از زنجیرههای هیدروکربنی بلند تشکیل شده میشوند. در این مطلب به این پرسش پاسخ میدهیم که تفاوت لیپید و چربی چیست.


لیپیدها ترکیبات آلی آبگریز و نامحلول در آب هستند که بیشتر آنها از اتصال زنجیرههای هیدروکربنی به نام اسید چرب و الکل مختلف تشکیل میشوند. چربی یکی از انواع لیپید و مولکول ذخیره انرژی در بعضی باکتریها، گیاهان و جانوران است. موم، گلیسروفسفولیپید، اسفنگولیپید، استروئید و لیپوپروتئين سایر لیپیدها هستند. موم در محافظت از ساختارهای زیستی، فسفولیپیدها در ساختار غشا، استروئیدها در انتقال پیام و لیپووپروتئینها در انتقال لیپیدها نقش دارند. در این مطلب از مجله فرادرس توضیح میدهیم تفاوت لیپید و چربی چیست و انواع لیپید چگونه ساخته میشود.
تفاوت لیپید و چربی چیست؟
لیپیدها گروه بزرگی از مولکولهای آلی آبگریز و نامحلول در آب هستند که در محلولهای آلی ازجمله اتر و استن حل میشوند. در ساختار این ترکیبات تعداد زیادی کربن وجود دارد. لیپیدها عملکرد متفاوتی دارند. بعضی از لیپیدها در ساختار غشای پلاسمایی سلولهای پروکاریوتی و یوکاریوتی وجود دارد. بعضی از آنها منبع یکی از منابع تامین انرژی این سلولها است.
بعضی از لیپیدهای کوچک انتقالدهنده پیام بین سلولها هستند. چربی یا تریگلیسرید یکی از انواع لیپیدها است که از یک الکل و سه زنجیره هیدروکربنی تشکیل میشود. فسفولیپدها، استروئیدها، موم، لیپیدهای اتری و گلیکولیپید انواع دیگر لیپیدها هستند. تفاوت چربیها با سایر لیپیدها در جدول زیر خلاصه شده است.
| نام لیپید | اجزای ساختار | عملکرد |
| چربی | گلیسرول و ۳ اسید چرب | ذخیره انرژی |
| موم | الکل و اسید چرب بلند | محافظت از ساختارهای خارجی بدن موجودات زنده |
| گلیسروفسفولیپید | فسفوگلیسرول و ۲ اسید چرب | شرکت در ساختار غشاهای زیستی و انتقال پیامهای سلولی |
| اسفنگولیپید | فسفواسفنگوزین و اسید چرب | شرکت در ساختار غشاهای زیستی و انتقال پیامهای سلولی |
| استروئید | هسته استروئیدی با چهار حلقه کربن | شرکت در سنتز غشای زیستی، پیشساز هورمونها و اسیدهای صفرا |
| لیپوپروتئین | چربی، کلسترول، فسفولیپید و پروتئین | انتقال لیپیدها در مایعات بدن |
ساختار چربی
چربیها یا تریگلیسریدها سادهترین انواع لیپیدها هستند که از یک گلیسرول و سه اسید چرب تشکیل شدهاند. گلیسرول الکلی با سه کربن و سه گروه هیدروکسیل است که هر گروه هیدروکسیل آن با گروه کربوکسیل یک اسیدچرب پیوند کوالانسی استر تشکیل میدهد. اسیدهای چرب زنجیره خطی از گروههای $$CH_2$$ با یک گروه کربوکسیل (COOH-) است. اسیدهای چرب را میتوان بر اساس ساختار به انواع اشباع و غیراشباع تقسیم کرد. تمام کربنهای اسیدچرب اشباع با پیوند یکگانه به هم متصل است و در زنجیره هیدروکربنی اسیدهای چرب غیراشباع حداقل یک پیوند دوگانه بین کربنها وجود دارد.
ساختار اسیدهای چرب غیراشباع سبب افزایش فاصله بین مولکولها و به مایع تبدیل شدن چربیها در دمای ۲۵ درجه سانتیگراد میشود. هیدروژن پیوند دوگانه بیشتر اسیدهای چرب طبیعی در یک طرف قرار دارد و کنفورماسیون سیس ایجاد میکند. این ساختار قبل از تجزیه در بدن به حالت ترانس تبدیل میشود و هیدروژنها در دو جهت مخالف قرار میگیرند.
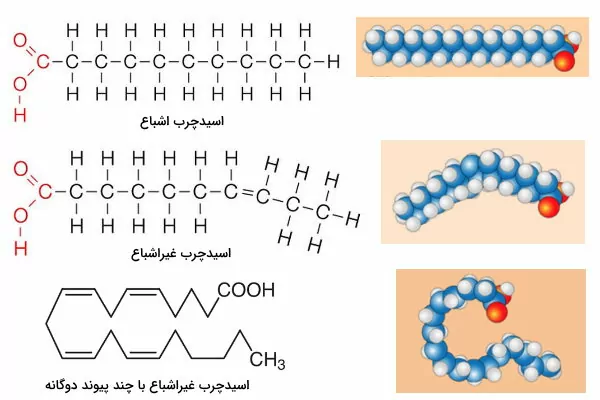
استفاده از نام منبعی که اسیدهای چرب برای اولین بار از آن استخراج شدهاند، یکی از روشهای نامگذاری این ترکیبات است. نام تعدادی از اسیدهای چرب همراه با تعداد کربنها، منبع اصلی، نامگذاری سیستمی و تعداد پیوندهای دوگانه در جدول زیر آمده است.
| نام | نام سیستمی | تعداد کربن و پیوند دوگانه | منبع اصلی |
| لائوریکاسید | n-دودکانوئیکاسید | ۱۲ – پیوند دوگانه ندارد. | روغن خرما |
| میریستیکاسید | n-تترادکانوئیکاسید | ۱۴ – پیوند دوگانه ندارد. | روغن خرما |
| پالمیتیکاسید | n-هگزادکانوئیکاسید | ۱۶ – پیوند دوگانه ندارد. | روغن زیتون |
| استئاریکاسید | n-اوکتادکانوسیکاسید | ۱۸ – پیوند دوگانه ندارد. | روغن نارگیل |
| بهنیکاسید | n-دوکوزانوئیکاسید | ۲۲ – پیوند دوگانه ندارد. | روغن تربچه |
| لیگنوسریکاسید | n-تتراکوزانوئیکاسید | ۲۴ – پیوند دوگانه ندارد. | موم درخت کارنوبا |
| پالمیتوئیکاسید | سیس-۹-هگزادکنوئیکاسید | ۱۶ – یک پیوند دوگانه دارد. | جلبک دریایی |
| اولئیکاسید | سیس-۹-اوکتادکنوئیکاسید | ۱۸ – یک پیوند دوگانه دارد. | روغن زیتون |
| لینولئیکاسید | سیس-۹، سیس-۱۲-اوکتادکا دیاِنوئیکاسید | ۱۸ – دو پیوند دوگانه دارد. | روغن ذرت |
| لینولنیکاسید | سیس-۹، سیس-۱۲، سیس-۱۵- اوکتادکا تریاِنوئیکاسید | ۱۸ – سه پیوند دوگانه دارد. | بافت جانوری |
| آراشیدونیکاسید | ۵،۸،۱۱،۱۴- ایکوزا تترا اِنوئیکاسید | ۲۰ – چهار پیوند دوگانه دارد. | بافت جانوری |
سه اسیدچرب متصل به گلیسرول ممکن است یکسان یا متفاوت باشند. طول زنجیره اسیدچرب در تریگلیسریدهای طبیعی متفاوت اما معمولا ۱۶، ۱۸ یا ۲۰ کربن است. تریگلیسریدهایی با اسیدهای چرب یکسان را میتوان بر اساس نوع اسیدچرب نامگذاری کرد. برای مثال تریاستئارین از سه استئاریکاسید، تریپلامیتین از سه پالمیتیکاسید و تریاولئین از سه اولئیکاسید تشکیل شده است. با وجود سر قطبی گلیسرول این ترکیبات در آب نامحلول هستند. در مسیرهای آنزیمی جانوران فقط اسیدهای چرب زوج کربن و خطی سنتز میشود. اما در باکتریها آنزیمهای سنتز اسیدهای چرب فرد کربن و شاخهدار وجود دارد.
تریگلیسرید منبع اصلی ذخیره انرژی در بیشتر سلولهای یوکاریوتی است. این ترکیب به شکل گرانولها سیتوپلاسمی در سلولها ذخیره میشود. تریگلیسرید مهرهداران در سلولهای تخصصیافته آدیپوسیت ذخیره میشود و در مواقع نیاز بدن به انرژی، آنزیم لیپاز این ترکیب را به اسید چرب و گلیسرول تجزیه میکند. گلیسرول ابتدا به گلیسرول ۳-فسفات و سپس به دیهیدروکسی استن فسفات تبدیل میشود. دیهیدروکسی استن فسفات در مسیر گلیکولیز به پیرووات تبدیل و در مسیر مشترک با کربوهیدراتها به سنتز ATP بیشتر کمک میکند. کربن اسیدهای چرب در میتوکندری، پراکسیزوم یا شبکه اندوپلاسمی به شکل استیل کوآنزیم از زنجیره جدا شده و با ورود به چرخه کربس در تامین ATP بیشتر شرکت میکند.
سنتز چربی ها
آنزیمهای سنتز چربیها تقریبا در تمام سلولهای جانوری وجود دارد. اما بیشتر مولکولهای چربی در کبد، سلولهای بافت چربی و سلولهای روده سنتز میشود. در سلولهای جانوری تریگلیسرید و گلیسروفسفولیپیدها از پیشسازهای گلیسرول ۳-فسفات و دیهیدروکسی استن فسفات در سلولهای کبدی و کلیه، و از مونوآسیل گلیسرول در سلولهای روده سنتز میشود. به علاوه بخشی از گلیسرول ۳-فسفات بافت چربی از در مسیر گلیکونئوژنز از پیروات تامین میشود. بخش زیادی از گلیسرول ۳-فسفات از تغییر ساختار دیهیدروکسی استن فسفات بهوسیله آنزیم ایزومراز و بخش کمی از آن با اضافه شدن گروههای فسفات بهوسیله آنزیم کیناز در سیتوپلاسم سنتز میشود.

مراحل بعدی سنتز در شبکه اندوپلاسمی ادامه مییابد. در مرحله اول آنزیم آسیل ترانسفراز یک اسید چرب به کربن ۱ گلیسرول و در مرحله بعدی یک اسید چرب به کربن ۲ اضافه میکند و فسفاتیدیکاسید سنتز تشکیل میشود. برای سنتز چربی، فسفاتیدیکاسید بهوسیله آنزیم فسفاتاز هیدرولیز و به ۱،۲-دیآسیل گلیسرول تبدیل میشود. در مرحله بعد با اضافه شدن اسید چرب سوم، تریگلیسرید سنتز میشود.
تفاوت چربی و موم
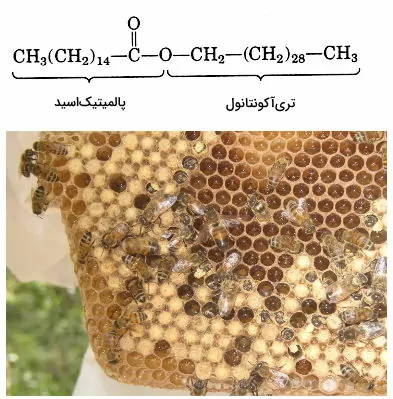
اگر تا این بخش از مطلب مجله فرادرس با ما همراهی کرده باشید، با ساختار و بیوسنتز چربیها آشنا شدهاید. در این بخش و بخشهای بعدی تفاوت انواع لیپید و چربی را توضیح میدهیم. مومها گروهی از لیپیدها هستند که مثل چربیها از اتصال اسیدچرب به الکل تشکیل میشوند. اما الکل این ترکیبات زنجیره هیدروکربنی بسیار بلندی (۱۶ تا ۳۰ کربن) است که با پیوند استری به یک اسیدچرب بلند (۱۴ تا ۳۶ کربن) اشباع یا غیراشباع متصل است. نقطه ذوب این ترکیبات از تریگلیسریدها بیشتر است و در دمای ۲۵ درجه سانتیگراد جامد هستند. نقش اصلی مومها محافظت از ساختارهای زیستی است. این ترکیبات آبگریز سطح برگ بعضی از گیاهان، پر پرندگان آبزی و کوتیکول بعضی از حشرات را میپوشاند. سطح مومی تبخیر آب از برگ گیاهان را کاهش میدهد و از خیس شدن بال پرندگان جلوگیری میکند. به علاوه موم منبع اصلی ذخیره انرژی در پلانکتونها است.
تفاوت چربی و فسفولیپید
فسفولیپیدها مثل چربی و موم از اتصال الکل به اسیدهای چرب تشکیل میشود. اما الکل این ترکیبات گلیسرول یا اسفنگوزین است که یکی از گروههای هیدروکسیل آن با فسفات جایگزین شده است. این ترکیبات از یک سر قطبی (فسفوالکل) و دم آبگریز (اسید چرب) تشکیل شدهاند. فسفولیپیدها را بر اساس نوع الکل به انواع گلیسروفسفولیپید و اسفنگولیپید تقسیم میکنیم. گلیسروفسفولیپیدها از اتصال دو اسید چرب به فسفوگلیسرول و اسفنگولیپیدها از اتصال یک اسید چرب به فسفواسفنگوزین تشکیل میشود.
گلیسروفسفولیپیدها، درشتمولکولهای اصلی ساختار غشای سلولی است. سر قطبی این لیپیدها دو طرف خارجی غشای دولایه (در تماس با سیتوپلاسم و مایع خارج سلولی) و دم آبگریز آنها وسط قرار میگیرد. کربن ۱ و ۲ گلیسرول فسفولیپیدها با پیوند استری به اسید چرب و کربن ۳ آن به گروه فسفات متصل میشود. معمولا اسیدچرب ۱۶ یا ۱۸ کربنه اشباع به کربن ۱ و اسیدچرب ۱۸ یا ۲۰ کربنه غیرشباع به کربن ۲ متصل است. به ترکیب تشکیل شده از اتصال اسیدهای چرب به فسفوگلیسرول فسفاتیدیکاسید گفته میشود.
از اتصال اتانول آمین، کولین، سرین، گلیسرول، اینوزیتول بیش فسفات و فساتیدیل کولین به فسافاتیدیکاسید به ترتیب فسفاتیدیل اتانول آمین، فسفاتیدیل کولین، فسفاتیدیل سرین، فسفاتیدیل گلیسرول، فسفاتیدیل اینوزیتول ۴،۵-بیس فسفات و کاردیولیپین تشکیل میشود. گروه اتصالی به فسفاتیدیکاسید، فسفاتیدیل سرین و فسفاتیدیل کولین خنثی است و این لیپیدها در pH = ۷ یک بار منفی (بار الکتریکی فسفات) دارند. گروه اتصالی به فسفاتیدیل اینوزیتول بیس فسفات چهار بار منفی و کاردیولیپین ۲ بار منفی دارد.
اسفنگوزین الکلی با ۱۸ کربن، دو گروه هیدروکسیل (کربن ۱ و ۳) و یک گروه آمین (کربن ۲) است که با کربوکسیل اسیدچرب پیوند آمیدی برقرار میکند. به ترکیب تشکیل شده از اتصال یک اسیدچرب به اسفنگوزین در شبکه اندوپلاسمی صاف سرآمید گفته میشود. گروه OH کربن ۱ سرآمید به یکی از گروههای فسفوکولین (فسفواسفنگولیپید)، فسفاتیدیل اتانول آمین (فسفواسفنگولیپید)، مونوساکارید (گلیکواسفنگولیپید) یا زنجیره اولیگوساکاریدی (گلیکواسفنگولیپید) متصل میشود. گروههای قطبی در جسم گلژی به سرآمید اضافه میشود. اسفنگولیپیدها بخشی از غشای پلاسمایی را میسازند و در انتقال پیامهای بین سلولی شرکت میکنند.

تفاوت سنتز چربی و فسفولیپید
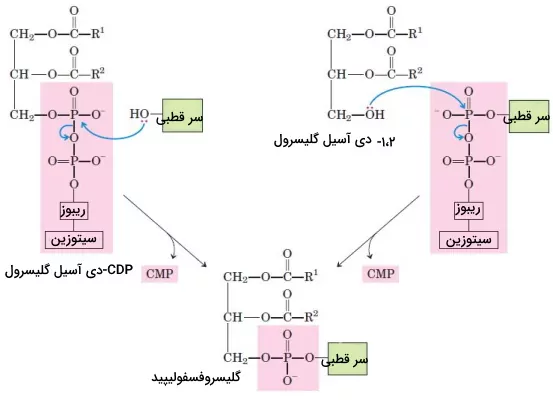
در بخش قبلی مطلب تفاوت لیپید و چربی توضیح دادیم که یکی از مکانیسمهای تشکیل فسفولیپیدها با تشکیل تریگلیسریدها مشترک است. در این بخش مکانیسم متفاوت سنتز فسفولیپیدها را بررسی میکنیم. در این مسیر گروه فسفات بهوسیله آنزیم کیناز به دیآسیل گلیسرول اضافه میشود. این آنزیم سر قطبی فسفولیپیدها را با پیوند فسفودیاستر به اسکلت کربنی الکل متصل میکند. در مرحله اول این فرایند هیدروکسیل آزاد دیآسیل گلیسرول یا سر قطبی به نوکلئوتید سیتیدین دیفسفات متصل میشود. با حمله نوکلئوفیلی هیدروکسیل مولکول دیگر سیتوزین مونوفسفات جدا و بین سرقطبی و اسکلت کربنی پیوند ایجاد میشود.
تفاوت چربی و استرول
ساختار استرولها تفاوت زیادی با ساختار تریگلیسریدها دارد. ساختار اصلی این ترکیبات از چهار حلقه با ۱۷ اتم کربن و یک گروه هیدروکسیل تشکیل شده است. تفاوت انواع استرول در گروهای عاملی متصل به حلقهها است. آنزیم سنتز این ترکیبات در سلولهای پروکاریوتی، گیاهان جانوران وجود دارد.
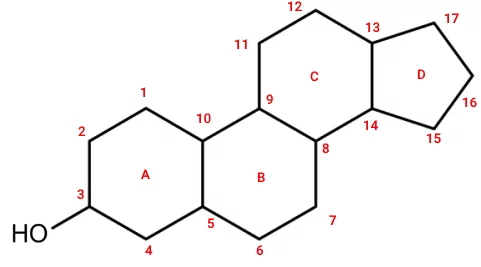
کلسترول بیشترین استرول جانوری است که درصد کمی از مولکولهای غشای پلاسمایی را به خود اختصاص میدهد و پیشساز سایر استروئیدها است. هورمونهای استروئیدی از تغییرات کلسترول در واکنشهای آنزیمی سنتز میشوند و فعالیتهای دستگاههای مختلف را تنظیم میکنند. این هورمونها برخلاف هورمونهای پپتیدی به راحتی با انتشار ساده از غشای پلاسمایی عبور میکنند و به گیرندههای سیتوپلاسمی یا هستهای متصل میشوند. اتصال این هورمونها به گیرنده با تغییر بیان ژن و عملکرد سلول همراه است.
قشر غدد فوق کلیه و غدد جنسی محل سنتز این هورمونها است. هورمونهای فوق کلیه به دو گروه گلوکوکورتیکوئید و مینروکورتیکوئیدها تقسیم میشوند. سنتز این هورمونها در شبکه اندوپلاسمی شروع و در میتوکندری کامل میشود. کورتیزول و کورتیکواسترون گلوکوکورتیکوئیدهایی هستند که متابولیسم بدن ازجمله سنتز گلوکز از آمینواسید و اسیدهای چرب، و سنتز گلیکوژن از گلوکز در کبد را کنترل میکند. به علاوه این هورمونها به تنظیم فشار خون و کاهش التهاب کمک میکنند. مینروکورتیکوئیدها ازجمله آلدوسترون غلظت نمکها و آب مایعات بدن را کنترل میکنند. هورمونهای جنسی بر اساس اسکلت کربنی به استروژنها با ۱۸ اتم کربن، اندروژنها با ۱۹ اتم کربن و پروژسترون با ۲۱ اتم کربن تقسیم میشوند. این هورمونها در رشد اندامهای جنسی و ایجاد صفات جنسی ثانویه نقش دارند.
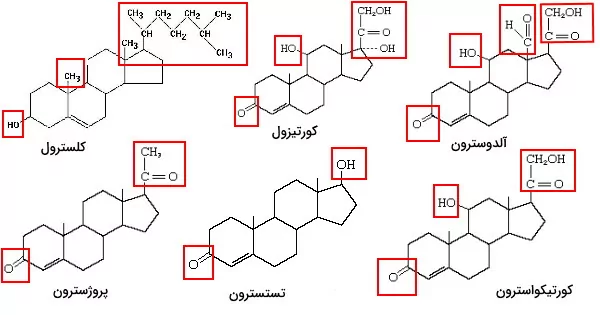
اسیدهای صفرا گروه دیگر استروئیدهای پستانداران هستند که در کبد از کلسترول سنتز میشوند. نمکهای صفرا از اتصال این اسیدها به تائورین یا گلایسین تشکیل میشود. این ترکیبات به گوارش چربیها در روده کوچک کمک میکنند.
تفاوت سنتز چربی و استرول
کلسترول پیشساز لیپیدهای استروئیدی در جانوران است. این لیپید ۲۷ کربنه از ترکیب مولکولهای استات در چهار مرحله سنتز میشود. در مرحله اول از ترکیب سه مولکول استیل کوآنزیم A، بتا-هیدروکسی-بتا متیلگلوتاریل-کوآ تشکیل میشود و آنزیم ردوکتاز این ترکیب را به موالونات تبدیل میکند. در مرحله بعدی موالونات به دو ایزوپرن فعال شکسته میشود. در این مرحله سه گروه فسفات از ۳ مولکول ATP به کربن ۳ و ۵ موالونات اضافه میشود. آنزیم دکربوکسیلاز با جدا کردن گروه کربوکسیل و گروه فسفات کربن ۳، ۳-فسفو-۵-پیروفسفوموالونات را به ایزوپنتیل پیروفسفات تبدیل میکند. آنزیم ایزومراز ایزوپنتیل پیروفسفات را به دی متیل آلیل پیروفسفات تغییر میدهد. در نتیجه دو ایزوپرن فعال برای ادامه سنتز کلسترول تشکیل میشود.
در مرحله بعدی، سر فسفات یکی از ایزوپرنها به انتهای ایزوپرن دیگر متصل و جرانیل پیروفسفات با ده اتم کربن سنتز میشود. جرانیل پیروفسفات به سر یک ایزوپرن دیگر ترکیب و فرانسیل پیروفسفات با ۱۵ کربن تشکیل میشود. در نهایت دو فرنسیل پیروفسفات با هم ترکیب و اسکوئالن تشکیل میشود. در مرحله آخر سنتز کلسترول، چهار حلقه هسته استروئیدی تشکیل میشود. برای تشکیل حلقه آنزیم اسکوئالن مونواکسیژناز یک اتم اکسیژن به انتهای این مولکول خطی اضافه میکند و اپوکسید تشکیل میدهد. این اپوکسید خطی با چند واکنش هماهنگ به ساختاری با چهار حلقه کربنی به نام لانوسترول و لانوسترول پس از ۲۰ مرحله واکنش آنزیمی به کلسترول تبدیل میشود.
تفاوت چربی و لیپوپروتئین
اگر تا این بخش از مطلب تفاوت لیپید و چربی با ما همراهی کرده باشید، متوجه تفاوت چربی با فسفولیپیدها، استروئیدها و موم شدهاید. لیپوپروتئینها آخرین ساختارهای لیپیدی هستند که در این مطلب بررسی میکنیم. لیپوپروتئین مجموعهای از چربی، کلسترول، فسفولیپید و آپوپروتئینها است که لیپیدها را در خون بین اندامها منتقل میکند. تریگلیسرید و کلسترول در مرکز این ساختار قرار دارد و لایهای از فسفولیپیدها پوششی اطراف آن تشکیل میدهد. آپوپروتئین در سطح و بین مولکولهای فسفولیپید قرار میگیرد. این ترکیبات بر اساس اندازه، ترکیب لیپیدها و نوع آپوپروتئین به گروههای لیپوپروتئین با چگالی زیاد (HDL)، لیپوپروتئین با چگالی کم (LDL)، لیپوپروتئین با چگالی متوسط (IDL)، لیپوپروتئین با چگالی بسیار کم (VLDL) و کیلومیکرون تقسیم میشود.
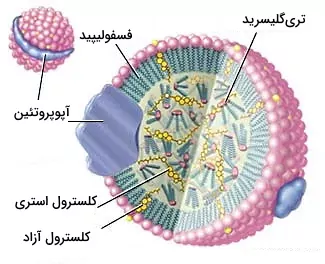
کیلومیکرونها لیپوپروتئینهایی هستند که تریگلیسرید غذا را از سلولهای دیواره روده به ماهیچه اسکلتی و آدیپوسیتها، و کلسترول غذا را به کبد منتقل میکنند. اندازه این لیپوپروتئينها بر اساس مقدار چربی غذا متفاوت است. آپوپروتئین B-48 پروتئین اصلی کیلومیکرونها است. تریگلیسرید این گلیکوپروتئین به وسیله آنزیمهای لیپاز در غشای ماهیچه اسکلتی و آدیپوسیت به اسیدهای چرب و گلیسرول تبدیل میشود. سلولهای ماهیچهای از تجزیه اسیدهای چرب برای سنتز ATP بیشتر استفاده میکنند. اسیدچرب در آدیپوسیتها به شکل چربی ذخیره میشود. باقیمانده کیلومیکرون در خون به کبد منتقل و با اندوسیتوز وارد سلولها میشود.
لیپوپروتئینهای با چگالی بسیار کم در کبد و از مقدار زیادی تریگلیسرید تشکیل میشود. پروتئین اصلی این ذرات آپوپروتئین B-100 و اندازه آنها بر اساس مقدار تریگلیسرید متفاوت است. تعداد تریگلیسرید این لیپوپروتئینها از کیلومیکرون کمتر و اندازه آنها کوچکتر است. اما تعداد کلسترول و فسفولیپید آن بیشتر است. جدا شدن تریگلیسرید در بافت ماهیچهای و آدیپوسیتها VLDL را به لیپوپروتئین یا چگالی متوسط تبدیل میکند. آپوپروتئین IDL مثل لیپوپروتئین با چگالی بسیار پایین B-100 است.
لیپوپروتئین با چگالی پایین از تغییر ترکیب لیپیدها در VLDL و IDL تولید میشود. مقدار کلسترول این ذرات از VLDL و IDL بسیار بیشتر و مقدار تریگلیسرید آن بسیار کمتر، اما آپوپروتئین آن B-100 است. بخش اصلی کلسترول خون در LDL وجود دارد. اتصال آپوپروتئین این ذرات به گیرنده LDL غشای سلولی با اندوسیتوز لیپوپروتئین همراه است. کلسترول لیپوپروتئینها بهوسیله آنزیمهای لیپاز لیزوزوم جدا میشود. لیپوپروتئین با چگالی بالا کمترین مقدار تریگلیسرید را دارد و مقدار کلسترول آن از IDL و LDL کمتر است. این ذرات کلسترول را از سلولهای مختلف به کبد منتقل میکنند. آپوپروتئین A-I، پروتئین اصلی HDL است.
سوالات متداول پیرامون تفاوت لیپید و چربی
در این بخش از مطلب مجله فرادرس به تعدادی از سوالات متداول پیرامون تفاوت لیپید و چربی پاسخ میدهیم.
تفاوت لیپیدهای ساده و مرکب چیست؟
لیپیدهای ساده از ترکیب اسیدهای چرب و الکل، و لیپیدهای مرکب از اضافه شدن کربوهیدرات، الکل دوم یا گروههای آمینی به اسکلت اصلی تشکیل میشوند.
تفاوت چربی و روغن چیست؟
به لیپیدهایی که در دمای ۲۵ درجه سانتیگراد جامد هستند چربی و به لیپیدهایی که در این دما مایع هستند، روغن گفته میشود.
source