موجودات یک گونه صفات مشترکی دارند. این صفات بهوسیله ماده ژنتیکی بین موجودات منتقل میشود. ماده ژنتیکی تمام موجودات زنده مولکولهای DNA است. مولکولهای DNA در پروکاریوتها و یوکاریوتها از دو رشته نوکلئوتیدی تشکیل شده است. اما ساختار آن متفاوت است. DNA بیشتر پروکاریوتها حلقوی و DNA بیشتر یوکاریوتها ازجمله انسان خطی است. نوکلئوتیدهای DNA در واحدهایی به نام ژن تقسیمبندی میشوند. هر ژن اطلاعات مربوط به یک پروتئین، tRNA یا rRNA را کد میکند. در فرایند رونویسی یک نسخه RNA از ژنها تولید میشود و در فرایند ترجمه کدهای ژنتیکی RNA به پروتئین ترجمه میشود. به علم بررسی این فرایندها ژنتیک گفته میشود. در این مطلب توضیح میدهیم ساختار DNA و RNA و فرایندهای مولکولی مربوط به آنها در ژنتیک چیست.
ژنتیک علم بررسی انتقال صفات بین نسلها است. در این علم تغییرات ژنتیکی که منجر به ایجاد تنوع بین افراد یک گونه یا ایجاد بیماریهای مختلف میشود را بررسی میکنیم. بهعلاوه با مطالعه ژنها میتوان روشهای درمانی جدید و بر اساس ویژگیهای فردی ایجاد کرد. در ابتدای این مطلب از مجله فرادرس ساختار مولکولهای ژنتیکی را مرور میکنیم. در بخشهای بعدی نحوه بیان ژن و تولید پروتئین را بررسی میکنیم. در ادامه روابط بین اللهای ژنی را مرور میکنیم و در آخر توضیح میدهیم گرایشهای علم ژنتیک چیست.
ژنتیک چیست؟
ژنتیک علم مطالعه انتقال صفات از یک نسل به نسل دیگر است. در این علم چگونگی به ارث رسیدن ژنها، ارتباط ژنها با صفات ظاهری، ارتباط ژنها با واکنشهای بیوشیمیایی بدن، ارتباط ژنها با هم، چگونگی تغییر ژنها، اثر محیط بر ژنها و ارتباط ژنها با بیماریها بررسی میشود. برای مثال به کمک مطالعات ژنتیکی میتوان مشخص کرد چرا رنگ موی یکی از دخترهای خانواده مشکی و دختر دیگری قهوهای است، چرا بعضی از بیماریها از والدین به فرزندان منتقل میشود، چرا بعضی از بیماریها فقط در مردان یک خانواده ایجاد میشود، چرا والدین سالم اما فرزندان آنها بیمار هستند، چرا رنگ گلها در نبود نور تغییر میکند یا چرا رشد باکتریها در محیطهای مختلف تغییر میکند. این سوالات با مطالعه ماده ژنتیکی سلولهای پروکاریوتی و یوکاریوتی پاسخ داده میشود.
ماده ژنتیکی تمام موجودات زنده مولکول DNA است. اطلاعات وراثتی به شکل کدهای ژنتیکی در این مولکول ذخیره شده است. در فرایند رونویسی نسخهای از این کدها در مولکولهای RNA کپی میشود و در فرایند ترجمه ژنتیکی به توالی آمینواسیدهای رشته پروتئینی ترجمه میشود. پروتئینها مولکولهایی هستند که در ایجاد صفات ظاهری و تغییر واکنشهای بیوشیمیایی نقش دارند. تغییر ژنها ساختار یا عملکرد این مولکولهای زیستی را تغییر میدهد. این تغییرات ممکن است با ایجاد صفات جدید همراه شود و صفت جدید ممکن است یک بیماری باشد.
علم ژنتیک به شاخههای ژنتیک مولکولی، اپیژنتیک، سیتوژنتیک، ژنتیک جمعیت و مهندسی ژنتیک تقسیم میشود. در مطلب ژنتیک چیست ابتدا ساختارها و فرایندهای مولکولی ژنتیک را بررسی میکنیم. در ادامه چگونگی به ارث رسیدن صفات را توضیح مرور میکنیم و در انتها توضیح میدهیم شاخههای علم ژنتیک چیست.
DNA چیست؟
در بخش قبلی مطلب ژنتیک چیست اشاره کردیم که ماده ژنتیکی موجودات زنده DNA است. در این بخش ساختار این درشتمولکول را بررسی میکنیم. ساختار مولکول DNA پلیمری دورشتهای است که از مونومرهای نوکلئوتیدی تشکیل شده است. هر نوکلئوتید DNA از یک باز آلی، یک قند دئوکسی ریبوز و گروه فسفات تشکیل میشود. کربن $$5^prime$$ قند دئوکسی ریبوز هر نوکلئوتید با پیوند کوالانسی به یک گروه فسفات و کربن $$1^prime$$ آن با پیوند کوالانسی به باز آلی متصل است. تیمین (T)، آدنین (A)، سیتوزین (C) و گوانین (G) بازهای آلی نوکلئوتیدهای هستند که در ساختار آنها یک یا دو حلقه نیتروژنی وجود دارد. آدنین و گوانین بازهای پورین با دو حلقه و تیمین و سیتوزین بازهای پیریمیدین با یک حلقه هستند. بازهای آلی دو رشته DNA با هم مکمل هستد و بین آنها پیوند هیدروژنی وجود دارد. باز A مکمل باز T و باز G مکمل باز C است. A و T دو و G و C سه پیوند هیدروژنی تشکیل میدهند.
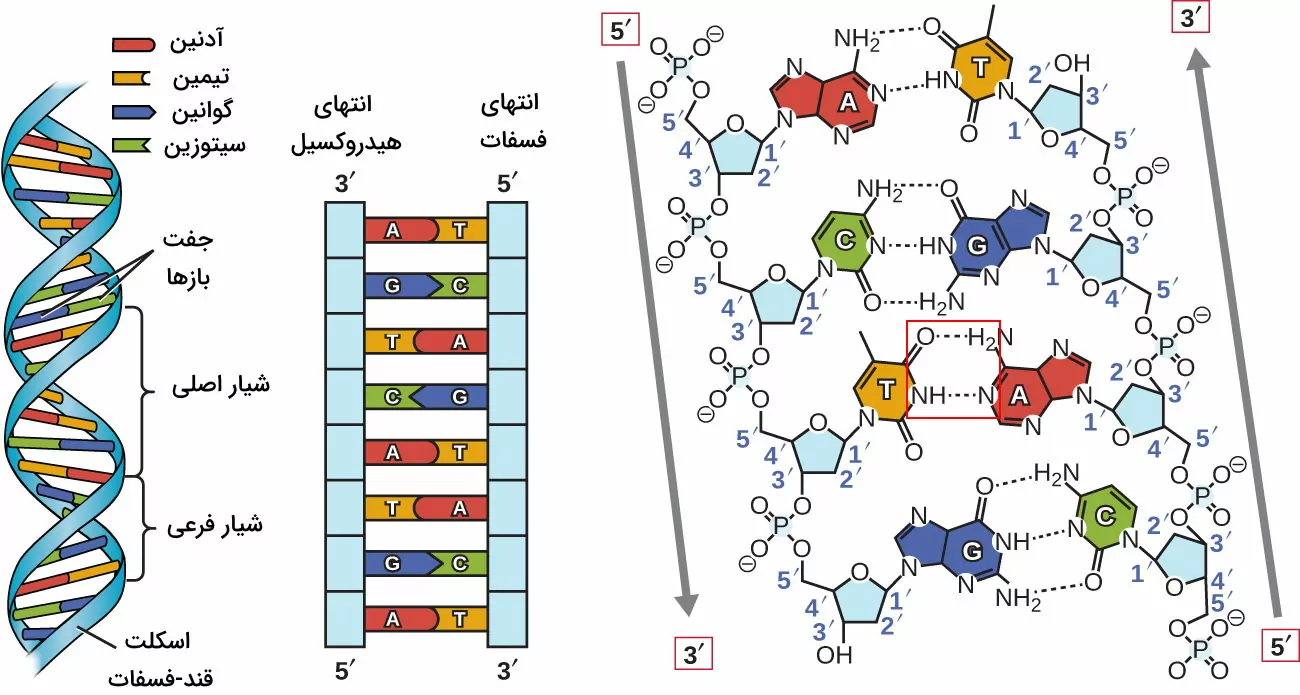
نوکلئوتیدها برای تشکیل رشتههای DNA با پیوند فسفودیاستر به هم متصل میشوند. در هر پیوند فسفودیاستر گروه هیدروکسیل (OH) متصل به کربن $$3^prime$$ یک نوکلئوتید به گروه فسفات نوکلئوتید بعدی متصل میشود. دو رشته DNA موازی و در جهت مخالف کنار هم قرار میگیرند. به این ترتیب انتهای فسفات (انتهای $$5^prime$$) یک رشته مقابل انتهای هیدروکسیل (انتهای $$3^prime$$) رشته دیگر قرار دارد. این دو رشته در جهت خلاف عقربههای ساعت دور یک محور فرضی میچرخند و ساختار مارپیچ دوگانه ایجاد میکنند. در هر مولکول DNA تعداد زیادی شیار اصلی و فرعی وجود دارد. شیار اصلی زمانی ایجاد میشود که دو رشته از هم دور میشوند و شیار فرعی زمانی ایجاد میشود که دو رشته به هم نزدیک میشوند. در هر پیچ DNA تقریبا ۱۰ جفت باز وجود دارد و فاصله بین پیچها تقریبا ۳٫۴ نانومتر است. قطر مولکول DNA تقریبا ۱٫۹ یا ۲۰ آنگستروم است. این مدل مولکولی اولین بار توسط واتسون و کریک ارائه شد و به آن ساختار B گفته میشود.
همانندسازی DNA
همانند سازی DNA فرایندی است که در آن از هر رشته DNA یک رشته مکمل ساخته میشود. در نتیجه در پایان هر همانندسازی دو مولکول دو رشته تشکیل میشود که کاملا شبیه هم و DNA والدی هستند. هر مولکول DNA جدید از یک رشته والدی و یک رشته تازه سنتز شده تشکیل شده است. به همین به این فرایند همانندسازی نیمه محافظتی نیز گفته میشود. DNA در تمام سلولها در جهت $$5^prime$$ به $$3^prime$$ همانندسازی میشود. به این ترتیب گروه فسفات نوکلئوتیدهای جدید به گروه هیدروکسیل نوکلئوتیدهای قبلی اضافه میشود.
در همانندسازی به رشته $$3^prime$$ به رشته $$5^prime$$، رشته «پیشرو» (Leading) و به رشته $$5^prime$$ به $$3^prime$$ «رشته پیرو» (Lagging) گفته میشود. رشته جدیدی که از پیشرو همانندسازی میشود $$5^prime$$ به $$3^prime$$ است. به همین دلیل در هر چنگال همانندسازی آنزیمها به راحتی نوکلئوتیدهای جدید را به انتهای هیدروکسیل نوکلئوتیدها اضافه میکنند و رشته جدید به شکل یک قطعه کامل همانندسازی میشود. اما رشته جدیدی که از پیرو همانندسازی میشود $$5^prime$$ به $$3^prime$$ است. آنزیمها رشته پیرو را به شکل قطعات کوچکی همانندسازی میکنند که در نهایت به هم متصل میشود و به آنها قطعات اوکازاکی گفته میشود. هر دور همانندسازی در سه مرحله شروع همانندسازی، طویلسازی DNA و خاتمه همانندسازی انجام میشود که در ادامه مطلب ژنتیک چیست توضیح میدهیم.
آنزیم های همانندسازی
DNA پلیمراز، هلیکاز، پرایماز، توپوایزومراز و پروتئینهای اتصالی به DNA تکرشتهای (SSB)، آنزیمها و پروتئینهای فرایند همانندسازی هستند.
- DNA پلیمراز: DNA پلیمراز آنزیمی است که رشته والد را الگو قرار میدهد و نوکلئوتیدها را برای سنتز رشته جدید به هم وصل میکند. در پروکاریوتها آنزیمهای پلیمراز I و II و II، و در یوکاریوتها آنزیمهای پلیمراز $$alpha$$ و $$beta$$ و $$gamma$$ و $$delta$$ و $$epsilon$$ در همانندسازی DNA شرکت میکنند.
- DNA پلیمراز I: این آنزیم فعالیت اگزونوکلئازی $$5^prime$$ به $$3^prime$$ و $$3^prime$$ به $$5^prime$$ دارد. این آنزیم با هیدرولیز پیوند فسفودیاستر، نوکلئوتیدها را از انتهای $$5^prime$$ و $$3^prime$$ رشتههای DNA جدا میکند. DNA پلیمراز I در فرایند ترمیم DNA باکتری شرکت میکند.
- DNA پلیمراز II: این آنزیم فعالیت پلیمرازی $$5^prime$$ به $$3^prime$$ و فعالیت اگزونوکلئازی $$3^prime$$ به $$5^prime$$ دارد.
- DNA پلیمراز III: پلیمراز III آنزیم اصلی همانندسازی DNA در باکتریها است که فعالیت پلیمرازی $$5^prime$$ به $$3^prime$$ و فعالیت اگزونوکلئازی $$3^prime$$ به $$5^prime$$ دارد.
- DNA پلیمراز $$alpha$$: این آنزیم در فعالیت اگزونوکلئازی $$3^prime$$ به $$5^prime$$ و فعالیت $$5^prime$$ به $$3^prime$$ پلیمرازی دارد. DNA پلیمراز $$alpha$$ در ترمیم DNA یوکاریوتی شرکت میکند.
- DNA پلیمراز $$beta$$: این آنزیم در فرایند ترمیم DNA شرکت میکند و فعالیت پلیمرازی $$5^prime$$ به $$3^prime$$ دارد.
- DNA پلیمراز $$gamma$$: این آنزیم DNA میتوکندری را همانندسازی میکند. DNA پلیمراز $$gamma$$ فعالیت پلیمرازی $$5^prime$$ به $$3^prime$$ و فعالیت اگزونوکلئازی $$3^prime$$ به $$5^prime$$ دارد.
- DNA پلیمراز $$delta$$: پلیمراز $$delta$$ رشته پیرو را همانندسازی میکند. این آنزیم فعالیت $$5^prime$$ به $$3^prime$$ پلیمرازی و فعالیت $$3^prime$$ به $$5^prime$$ اگزونوکلئازی دراد.
- DNA پلیمراز $$epsilon$$: پلیمراز $$epsilon$$ آنزیم همانندسازی رشته پیشرو DNA یوکاریوتی است که فعالیت پلیمرازی $$5^prime$$ به $$3^prime$$ و فعالیت اگزونوکلئازی $$5^prime$$ به$$3^prime$$ دارد.
- هلیکاز: هلیکاز موتورپروتئین آنزیمی است که جلوی آنزیمهای پلیمراز حرکت و با هیدرولیز پیوند هیدروژنی دو رشته DNA را از هم جدا میکند.
- پرایماز: آنزیمهای پلیمراز برای شروع همانندسازی DNA به انتهای OH آزاد نیاز دارند. آنزیمهای پرایماز قطعه RNA چند نوکلئوتیدی مکمل رشته الگو به نام پرایمر میسازد و پلیمراز همانندسازی را از انتهای OH آزاد ریبونوکلئوتیدها شروع میکند.
- توپوایزومراز: آنزیمهای توپوایزومراز پیچهای اضافه ایجاد شده در DNA حین همانندسازی رونویسی را از بین میبرند. این آنزیمها پیوند فسفودیاستر بین نوکلئوتیدها را در یک یا دو رشته هیدرولیز میکنند. توپوایزومرازهای I در یک رشته DNA و توپوایزومرازهای II در دو رشته DNA شکست ایجاد میکند. توپوایزامرازهای II در چرخه سلولی و فشروده شدن کروموزومها نقش مهمی دارند. DNA جیراز، توپوایزومراز II باکتریها است.
- پروتئینهای اتصالی به DNA تکرشتهای: این پروتئینها به DNA تکرشتهای متصل میشوند و از به هم متصل شدن DNA در حین همانندسازی و تجزیه DNA بهوسیله آنزیمهای نوکلئاز جلوگیری میکنند.
شروع همانندسازی
در DNA باکتری یک و در DNA یوکاریوتها چند جایگاه شروع رونویسی وجود دارد. این جایگاه از نوکلئوتیدهای A و T تشکیل شده است که باز کردن پیوند هیدروژنی آنها به انرژی کمتری نیاز دارد. فاکتورهای شروع این جایگاه را شناسایی میکنند. هلیکاز یکی از این پروتئینها است که دو رشته DNA را از هم باز و ساختاری شبیه Y ایجاد میکند که به آن چنگال همانندسازی گفته میشود. پس از باز شدن دو رشته پروتئینهای SSB به رشتههای DNA متصل میشوند و پرایمر سنتز RNA پرایمر را شروع میکند.

طویل سازی DNA و خاتمه همانندسازی
پس از سنتز پرایمر آنزیم DNA پلیمر همانندسازی رشته های الگو را در دو طرف چنگال همانندسازی شروع میکند. پس از کامل شدن همانندسازی DNA چنگال همانندسازی، آنزیمهای پلیمراز با فعالیت اگزونوکلئازی پرایمر را تجزیه و با فعالیت پلیمرازی دئوکسی ریبونوکلئوتیدهای جدید را جایگزین آن میکند. در آخر آنزیم لیگاز آخرین نوکلئوتید رشته جدید و اولین نوکلئوتید رشتهای که جایگزین پرایمر شده، پیوند ایجاد میکند.
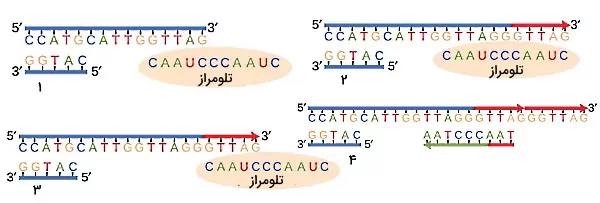
در DNA خطی یوکاریوتها زمانی که آخرین پرایمر انتهای $$3^prime$$ رشته پیرو برداشته میشود، انتهای OH آزاد برای فعالیت آنزیم پلیمراز وجود ندارد. در نتیجه بخشی از DNA در انتهای این رشته همانندسازی نمیشود و ممکن است بخشی از اطلاعات وراثتی از بین برود. اما در انتهای DNA خطی انسان و سایر پستانداران تعداد زیادی توالی تکراری $$5^prime TTAAGGG3^prime$$ به نام تلومر وجود دارد که به همانندسازی انتهای DNA کمک میکند. DNA تلومر تکرشتهای است و بهوسیله آنزیم تلومراز همانندسازی میشود. در این آنزیم RNA وجود دارد که مکمل توالی تلومر است و الگوی تلومراز برای اضافه کردن نوکلئوتیدها به انتهای $$3^prime$$ رشته پیرو اضافه میکند. پس از اضافه شدن نوکلئوتیدها آنزیم پرایمراز و پلیمراز ناحیه انتهای رشته پیرو را همانندسازی میکنند.
کروموزوم چیست؟
پس از مرور ساختار DNA و نحوه سنتز آن در بخشهای قبلی، در این بخش از مطلب ژنتیک چیست ساختار کروموزوم را بررسی میکنیم. کروموزم DNAای فشرده شده بهوسیله پروتئینها است. تعداد و شکل کروموزومها در موجودات مختلف متفاوت است. کروموزوم باکتریها در ناحیه نوکلئوئیدی، کروموزوم سلولهای جانوری در هسته و میتوکندری و کروموزوم سلولهای گیاهی در هسته و کلروپلاست قرار دارد. ساختار کروموزوم هستهای خطی و ساختار کروموزوم باکتری، میتوکندری و کلروپلاست حلقوی است.
کروموزوم یوکاریوتی از ساختارهای تکراری به نام نوکلئوزوم تشکیل شده است. هر نوکلئوزوم مجموعهای از پروتئینهای اسیدی هیستون و DNA است.H1 و H2A و H2B و H3 و H4 پروتئینهای هیستون هستند که یک ساختار اوکتامری تشکیل میدهند. این ساختار از دو دیمر H2A|H2B و H3|H4 تشکیل شده است که ۱۴۶ جفت باز (تقریبا دو دور) DNA دور آن پیچیده است. هیستون H1 خارج از این ساختار اوکتامری به دو انتهای DNA متصل میشود و به افزایش پایداری نوکلئوزوم کمک میکند. نوکلئوزومها با توالی ۸۰ نوکلئوتیدی از هم جدا میشوند. تشکیل نوکلئوزومها DNA را به رشتههایی با قطر ۱۰ نانومتر تبدیل میکند که به آن کروماتین گفته میشود. از برهمکنش H1 پیچخوردگیهای بیشتری در کروماتین ایجاد و رشته ۳۰ نانومتری تشکیل میشود. از پیچخوردگی بیشتر کروماتین رشتههایس با قطر ۷۰۰ نانومتری و ۱۴۰۰ نانومتری ایجاد میشود. رشته ۱۴۰۰ نانومتری کروموزومهای مرحله متافاز تقسیم سلولی است که فشردهترین حالت DNA است.
هیستونها در کروموزوم باکتریها وجود ندارد. اما پروتئینهای HU و آنزیم توپوایزومراز I با تشکیل سوپرکویل (ابرمارپیچ) مثبت و منفی DNA این موجودات را فشرده میکنند. پروتئینهای HU ساختارهای تترامری تشکیل میدهند که DNA سوپرکویل دور آن میچرخد. سوپرکویلهای مثبت هم جهت مارپیچ DNA و سوپرکویلهای منفی خلاف جهت مارپیچ DNA هستند.
کروموزوم های انسان
در هر سلول سوماتیک انسان ۲۳ جفت کروموزوم با شکل و اندازه متفاوت وجود دارد. کروموزوم ۱ بلندترین و کروموزوم ۲۲ کوتاهترین کروموزوم این سلولها است. ۲۲ جفت این کروموزومها اوتوزومی و یک جفت آن جنسی است. به کروموزومهای جنسی X و Y گفته میشود که در تعیین جنسیت نقش دارند. در سلولهای جنس نر ۲۲ جفت کروموزوم اتوزومی، یک کروموزوم Y و یک کروموزوم X وجود دارد. در سلولهای جنس ماده ۲۲ جفت کروموزم اتوزومی و دو کروموزوم X وجود دارد. فرزندان یک جفت کروموزوم اتوزومی و یک کروموزوم جنسی از هر والد دریافت میکنند.

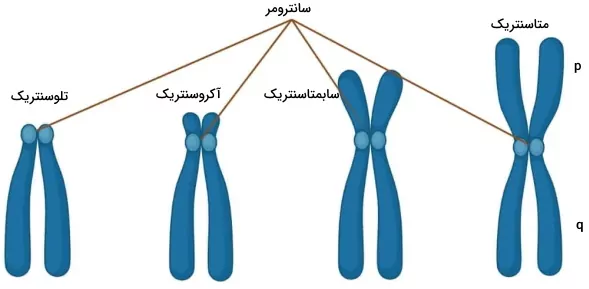
کروموزمهای متافازی از سانترومر، بازوی بلند، بازوی کوتاه و تلومر تشکیل شده است. سانترومر بخشی از DNA است که از توالیهای تکراری زیاد تشکیل شده است. توالی DNA در این بخش نقش ساختاری دارد و ژنی کد نمیکند. این بخش DNA محل اتصال دو کروموزوم همولوگ در متافاز میتوز و میوز به هم است. به قطعات DNA در دو طرف سانترومر بازوی بلند (p) و کوتاه (q) گفته میشود. کروموزومها بر اساس محل قرار گرفتن سانترومر به انواع متاسنتریک، سابمتاسنتریک، آکروسنتریک و تلوسنتریک تقسیم میشوند.
- کروموزوم متاسنتریک: سانترومر این کروموزومها در وسط قرار دارد و قطعات DNA دو طرف آن با هم برابر است. کروموزومهای ۱، ۳، ۱۶، ۱۹ و ۲۰ انسان متاسنتریک هستند.
- کروموزوم سابمتاسنتریک: سانترومر این کروموزومها نزدیک مرکز کروموزوم قرار دارد و یکی از بازوها کمی بلندتر از بازوی دیگر است. کروموزومهای ۲، ۴ تا ۱۲، ۱۷، ۱۸ و X سابمتاسنتریک هستند.
- کروموزوم آکروسنتریک: سانترومر این کروموزومها نزدیک انتهای کروموزوم قرار دارد واختلاف اندازه دو بازو بیشتر است. کروموزومهای ۱۳،۱۴، ۱۵، ۲۲ و Y آکروسنتریک هستند.
- کروموزوم تلوسنتریک: سانترومر این کروموزومها در یکی از دو انتهای کروموزوم قرار دارد. این نوع کروموزوم در سلولهای انسان وجود ندارد.
کروموزوم همولوگ و غیر همولوگ در ژنتیک چیست؟
کروموزومهای همولوگ به جفت کروموزومی گفته میشود که توالی ژنی، لوکوسهای ژنی، اندازه و محل قرار گرفتن سانترومر آنها یکسان است. تبادل ژنی بین این کروموزومها سبب ایجاد نوترکیبی میشود. در سلولهای جنس ماده انسان ۲۳ جفت کروموزوم همولوگ و در سلولهای نر ۲۲ جفت کروموزوم همولوگ وجود دارد (X و Y همولوگ نیستند). به این ترتیب کروموزوم ۱ که از پدر به ارث رسیده است با کروموزوم ۱ که از پدر به ارث رسیده است همولوگ و با کروموزومها ۲ غیرهمولوگ است. کروموزمهای غیرهمولوگ به جفت کروموزومی گفته میشود که توالی ژنی، لوکوسهای ژنی، اندازه و محل قرار گرفتن سانترومر متفاوتی دارند. تبادل ژنی بین این کروموزومها منجر به جهش ژنتیکی میشود.
هاپلوئید و دیپلوئید در ژنتیک چیست؟
در سلولهای هاپلوئید یک دسته، در سلولهای دیپلوئید دو دسته و در سلولهای پلیپلوئید بیش از دو دسته کروموزومهای همولوگ وجود دارد. بیشتر باکتریها یک کروموزوم دارند و هاپلوئید هستند. تنها سلولهای هاپلوئید بدن انسان گامتها (اسپرم و تخمک) و سایر سلولهای انسان دیپلوئید هستند. تعداد کروموزومهای همولوگ در هر دسته را با حرف n نشان میدهند و تعداد دستهها را قبل از آن مینویسند. به این ترتیب تعداد کروموزومهای انسان ۲n=۴۶ است.
ژن چیست؟
ژن قطعهای از DNA است که اطلاعات ژنتیکی را به نسل بعد منتقل میکند. بیشتر ژنها اطلاعات مربوط به سنتز پروتئینها را کد میکنند. به طور کلی هر ژن از یک ناحیه پروموتر، یک ناحیه کدکننده و ناحیه خاتمه تشکیل شده است. پروموتر ناحیه از ژن است که آنزیمها و پروتئینهای تنظیمی بیان ژن به آن متصل میشود. پروموتر در ابتدای ژن قرار دارد. ناحیه کدکننده، توالی است که کدهای ژنتیکی سنتز پروتئینها یا RNA را دارد. ناحیه خاتمه، توالی است که بیان ژن را تمام میکند. به توالیهای نزدیک انتهای $$5^prime$$ بالادست ژن و نواحی نزدیک $$3^prime$$ پایین دست ژن گفته میشود. ساختار ژن را همیشه در رشته $$5^prime$$ به $$3^prime$$ نشان میدهیم. ساختار ژن در پروکاریوتها و یوکاریوتها تفاوت دارد.
- پروکاریوتها: در سلولهای پروکاریوتی چند ناحیه کدکننده پروتئینهای متفاوت بهوسیله یک پروموتر کنترل میشود. پروموتر تمام ژنهای پروکاریوتی در نواحی ۱۰- (TATAAT) و ۳۵- (TTGACA) است. در هر ژن پروکاریوتی یک اوپراتور وجود دارد. اوپراتور توالی نزدیک پروموتر است که پروتئینهای تنظیمی به آن متصل میشود. به مجموعه پروموتر، اوپراتور و نواحی کدکننده آن یک اپران گفته میشود.
- یوکاریوتها: به ناحیه کدکننده ژن یوکاریوتها «چارچوب خوانش باز» (Open Reading Frame | ORF) گفته میشود که از توالیهای اگزون و اینترون تشکیل شده است. اگزونهای توالیهای کدکننده پروتئین و اینترونها نواحی ساختاری بین اگزونها هستند. پروموتر سلولهای یوکاریوتی از سه ناحیه بالادست، مرکزی و پاییندست قرار دارد. آنزیمها و پروتئینهای شروع رونویسی به پروموتور مرکزی و پروتئینهای تنظیم بیان ژن به پروموترهای بالادست و پایین دست متصل میشود. پروموتر مرکزی نزدیک جایگاه شروع رونویسی قرار دارد و آنزیمها به توالی $$5^prime TATAA 3^prime$$ آن درناحیهای به نام جعبه TATA متصل میشوند. به پرموتورهای بالادست و پاییندست افزاینده و کاهنده نیز گفته میشود. اتصال فاکتورهای تنظیمی به توالیهای کاهنده بیان ژن را افزایش و اتصال این پروتئینها به توالیهای کاهنده بیان ژن را کاهش میدهد. بین پروموتر و جایگاه شروع رونویسی و پس از آخرین اگزون توالی وجود دارد که رونویسی میشود اما ترجمه نمیشود. این توالی $$5^prime{UTR}$$ و $$3^prime{UTR}$$ نام دارد.
ژنوتیپ و فنوتیپ در ژنتیک چیست؟
ژنوتیپ نشاندهنده توالی DNA در جایگاه مشخصی از کروموزوم و فنوتیپ نشاندهنده ویژگیهای ظاهری، بیوشیمیایی و رفتاری ایجاد شده بهوسیله ژن است. برای مثال رنگ چشم انسان فنوتیپ و ژن تعیینکننده رنگ چشم ژنوتیپ است.
الل در ژنتیک چیست؟
اللها شکلهای متفاوت یک ژن هستند. بعضی از ژنها ۲ الل و بعضی از ژنها بیش از دو الل دارند. اگر اللهای ژن در یک فرد یکسان باشد، فرد برای آن ژن هموزیگوت یا خالص و اگر اللهای ژن در یک فرد متفاوت باشد، فرد برای آن ژن هتروزیگوت یا ناخالص است.
RNA چیست؟
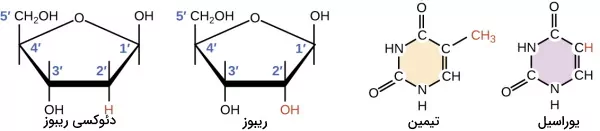
RNA نوکلئیکاسید دیگری است که از زیرواحدهای نوکلئوتیدی تشکیل شده است. نوکلئوتیدهای RNA از بازهای نیتروژنی، قند ریبوز و گروه فسفات تشکیل میشود. گوانین، سیتوزین، آدنین و یوراسیل بازهای نیتروژنی ریبونوکلئوتیدها هستند. در بخشهای قبلی این مطلب ژنتیک چیست از مجله فرادرس ساختار بازهای A و G و C را توضیح دادیم. یوراسیل بازی است که در ساختار RNA جایگزین تیمین شده است. تنها تفاوت یوراسیل و تیمین حذف گروه متیل متصل به حلقه در یوراسیل است. ریبوز قند پنجکربنه و تنها تفاوت ساختار آن با دئوکسی ریبوز اضافه شدن گروه هیدروکسیل (OH) به کربن $$2^prime$$ است. ریبونوکلئوتیدهای ساختار RNA با پیوند فسفودیاستر به هم متصل هستند.
مولکول RNA برخلاف DNA از یک رشته تشکیل شده است. این رشته در بعضی از انواع RNA تا میخورد و پیوندهای هیدروژنی درونمولکولی بین بازهای هیدروژنی ایجاد میشود. mRNA و rRNA و tRNA سه نوع اصلی RNA در سلولهای یوکاریوتی و پروکاریوتی هستند که در ترجمه نقش دارند.
- mRNA: مولکول mRNA یا RNA پیامرسان از رونویسی ژنهای کدکننده پروتئین تولید میشود. این RNA در پروکاریوتها پس از رونویسی بدون تغییر و در یوکاریوتها پس از تغییراتی به پروتئین ترجمه میشود. نوکلئوتیدهای mRNA به زیرواحدهای سهتایی به نام کدون تقسیم میشوند. هر کدون نشاندهنده یک آمینواسید است. برای مثال کدون AUG در پروتئین به آمینواسید متیونین ترجمه میشود.
- rRNA: مولکول rRNA یا RNA ریبوزومی در ساختار ریبوزومها شرکت میکند و از رونویسی توالیهای بلندی تولید میشود که به مولکولهای کوچکتر تقسیم میشوند. رونویسی و سرهمبندی این مولکولها با پروتئین در هسته سلولهای یوکاریوتی و سیتوپلاسم سلولهای پروکاریوتی انجام میشود. rRNA فعالیت آنزیمی دارد و تشکیل پیوند پپتیدی در فرایند ترجمه را کاتالیز میکند. ریبوزوم باکتریها از rRNAهای ۱۶S و ۲۳S و ۵S و ریبوزوم یوکاریوتها از rRNAهای ۱۸S و ۲۸S و ۵٫۸S تشکیل شده است.
- tRNA: مولکول tRNA یا RNA ناقل در ترجمه mRNA به پروتئین نقش دارد. ساختار دوبعدی این مولکولها شبیه برگ شبدر است و از یک انتهای $$3^prime$$ پذیرنده، انتهای $$5^prime$$ فسفات، بازوی D، بازوی T و بازوی آنتیکدون تشکیل شده است. نوکلئوتید A توالی CAA در انتهای $$3^prime$$ این مولکولها بهوسیله آنزیم پپتیدیل ترانسفراز به گروه کربوکسیل آمینواسید متصل میشود. بازوی آنتیکدون tRNA از ریبونوکلئوتیدهایی تشکیل شده است که با پیوند هیدروژنی به کدونهای mRNA متصل میشوند. هر tRNA به یک نوع آمینواسید متصل میشود و نوع آمینواسید را توالی آنتیکدون مشخص میکند.
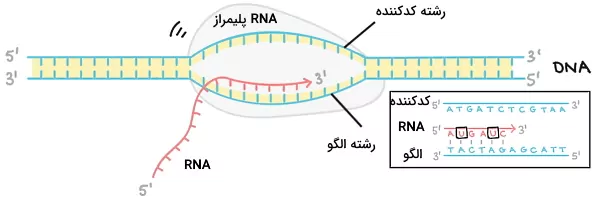
رونویسی ژنتیک چیست؟
اگر تا این بخش از مطلب ژنتیک چیست با ما همراهی کرده باشید، متوجه شدید که ماده ژن چیست و از چه بخشهایی تشکیل شده است. در این بخش توضیح میدهیم رونویسی در ژنتیک چیست. رونویسی فرایندی است که یک مولکول RNA از مولکول DNA ساخته میشود. آنزیمهای RNA پلیمراز رشته $$3^prime$$ به $$5^prime$$ مولکول DNA را الگو قرار میدهد و ریبونوکلئوتیدها را برای سنتز RNA به هم وصل میکنند. تمام ژنهای پروکاریوتی بهوسیله یک آنزیم پلیمراز رونویسی میشود. اما در سلولهای یوکاریوتی سه نوع RNA پلیمراز برای رونویسی ژنهای پروتئینی، rRNA و tRNA وجود دارد. این آنزیمها در سه مرحله شروع، طویلسازی و خاتمه DNA را رونویسی میکنند. این سه مرحله در پروکاریوتها و یوکاریوتها تفاوتهایی دارد که در ادامه مطلب ژنتیک چیست آنها را توضیح میدهیم.
رونویسی در پروکاریوت
پروکاریوتها هسته ندارند. به همین دلیل رونویسی و ترجمه در این سلولها همزمان و در سیتوپلاسم انجام میشود. RNA پلیمراز پروکاریوتها از دو زیرواحد آلفا ($$alpha$$)، یک زیرواحد سیگما ($$sigma$$)، یک زیرواحد بتا ($$beta$$) و یک زیرواحد بتا پریم ($$beta^prime$$) تشکیل شده است. زیرواحدهای آلفا به قرار گرفتن زیرواحدهای دیگر پلیمراز روی DNA کمک میکنند، زیرواحد بتا به دنباله RNA در حال سنتز متصل میشود، بتا پریم به مولکول DNA متصل میشود و زیرواحد سیگما فقط به شروع شدن رونویسی کمک میکند. زیرواحدهای آلفا، بتا و بتا پریم هسته آنزیمی را میسازند و زیرواحد سیگما بخش کمکی است که پس از شروع رونویسی جدا میشود. زیرواحد سیگما پروموتر را تشخیص میدهد. در نبود این زیرواحد رونویسی از نوکلئوتید شروع و پروتئین عملکردی تولید نمیشود.
سنتز رشته RNA پس از جدا شدن زیرواحد سیگما شروع میشود و تا رسیدن به توالی خاتمه ادامه دارد. در این مسیر پیوند هیدروژنی بازها بین دو رشته DNA در جلوی آنزیم باز و پس از رونویسی دوباره تشکیل میشود. رونویسی با روش وابسته به پروتئین Rho و تشکیل سنجاقسر خاتمه مییابد.
- روش وابسته به پروتئین Rho: در این روش پروتئین Rho پشت سر RNA پلیمراز روی DNA حرکت میکند. در انتهای ژن توالی از نوکلئوتیدهای GC وجود دارد که هیدرولیز پیوند هیدروژنی آن به انرژی و زمان بیشتر نیاز دارد. به همین دلیل RNA پلیمراز در انتهای ژن متوقف میشود. در این حالت برهمکنش پروتئین Rho با پلیمراز منجر به تغییر کنفورماسیون این آنزیم، جدا شدن آن از DNA و خاتمه رونویسی میشود.
- تشکیل سنجاقسر: در این روش پس از رونویسی توالی انتهایی غنی از GC، انتهای RNA خم میشود. در این حالت از اتصال بازهای G و C انتهایی ساختار سنجاق سری تشکیل میشود. این ساختار RNA پلیمراز را در ابتدای یک توالی غنی از AT متوقف میکند. تشکیل پیوند هیدروژنی ضعیف بین توالی UA رونویسی شده و توقف پلیمراز پایداری این آنزیم روی DNA را کاهش میدهد. در نتیجه آنزیم از DNA جدا میشود.
رونویسی در یوکاریوت
در بخشهای قبلی مطلب ژنتیک چیست توضیح دادیم که DNA یوکاریوتها بهوسیله پروتئينهای هیستون فشرده شده است. قبل از شروع رونویسی فشردگی DNA باید کمتر شود. اضافه شدن گروههای متیل و استیل بهوسیله آنزیمهای استیلاز و متیلاز به انتهای کربوکسیل هیستونها، بار مثبت هیستون را کاهش میدهد، برهمکنش هیستون با DNA کاهش مییابد و فضای کافی برای اتصال RNA پلیمراز به DNA ایجاد میشود. رونویسی یوکاریوتها در هسته و بهوسیله آنزیمهای پلیمراز I و II و III انجام میشود. RNA پلیمراز I ژنهای rRNA، آنزیم RNA پلیمراز II ژنهای پروتئین و RNA پلیمراز II ژنهای tRNA و rRNAهای بسیار کوچک (۵S RNA) را رونویسی میکند. در ادامه این مطلب رونویسی ژنهای پروتئینی بهوسیله آنزیم پلیمراز II را توضیح میدهیم. رونویسی این ژنها در چهار مرحله تشکیل کمپلکس پیش از رونویسی، شروع رونویسی، طویلسازی mRNA و خاتمه انجام میشود.
تشکیل کمپلکس پیش از رونویسی
برخلاف پروکاریوتها، آنزیم پلیمراز یوکاریوتی بهوسیله پروتئینهای کمکی به DNA متصل میشود. این پروتئینها بر اساس دو حرف اول انگلیسی کلمه «Transcription Factor» به معنی فاکتور رونویسی و حروف الفبای انگلیسی نامگذاری شده است. پروتئین TFIID اولین فاکتور رونویسی است که در شیار کوچک DNA به جعبه TATA متصل میشود. این پروتئین خمیدگی کوچکی در DNA ایجاد میکند. در مرحله بعد TFIIA به TFIID متصل میشود و پایداری کمپلکس ایجاد شده را افزایش میدهد. همزمان TFIIB به شیار بزرگ DNA متصل میشود. این پروتئین جهت رونویسی را تعیین میکند. در مرحله TFIIF آنزیم RNA پلیمراز II را به فاکتورهای رونویسی دیگر متصل میکند. پس از اتصال پلیمراز TFIIE پروتئین TFIIH را به کمپلکس اضافه میکند.
شروع رونویسی
TFIIH از یک زیرواحد هلیکاز، یک زیرواحد DNA ترانسلوکاز و یک زیرواحد کیناز تشکیل شده است. پس از تشکیل کمپلکس پیش از رونویسی زیرواحد ترانسلوکاز روی DNA حرکت میکند و آنزیم پلیمراز را با خود به جلو میکشد. فشار ایجاد شده سبب باز شدن دو رشته DNA از هم و قرار گرفتن DNA در جایگاه فعال آنزیم میشود. در این حالت توالی ۳ تا ۷ بازی از DNA رونویسی میشود. در ادامه زیرواحد کیناز TFHII به سرینهای انتهای کربوکسیل پلیمراز گروههای فسفات اضافه میکند، پلیمراز از فاکتورهای رونویسی جدا میشود و توالی ۱۴ تا ۲۲ نوکلئوتیدی DNA رونویسی میشود.
طویل سازی
پس از جدا شدن از فاکتورهای شروع رونویسی، پلیمراز روی DNA حرکت و ریبونوکلئوتیدها را در جهت $$5^prime$$ به $$3^prime$$ پشت سر هم اضافه میکند. در این مرحله پروتئین دیمر FACT در جلوی پلیمراز یکی از دیمرهای H2A|H2B را بر میدارد و پلیمراز DNA نوکلئوزومی را رونویسی میکند. پس از رونویسی قطعه DNA، نوکلئوزوم دوباره تشکیل میشود.
پایان رونویسی
رونویسی در ناحیه $$3^prime{UTR}$$ تمام و پلیمراز از DNA میشود. «فاکتور اختصاصی جداسازی و پلیآدنیلاسیون» (Cleavage and Polyadenylation Specificity Factor | CPSF)، فاکتور تحریککننده جداسازی (Cleavage Stimulating Factor | CSTF) و فاکتورهای جداسازی I و II پروتئین اصلی در پایان رونویسی DNA انسان هستند. در ناحیه $$3^prime{UTR}$$ یک ۶ تا ۸ نوکلئوتیدی پلی A وجود دارد. رونویسی این توالی اولین سیگنال خاتمه رونویسی است. CPSF و CSTF و فاکتورهای جداسازی I و II به توالی پلی A متصل میشود. این کمپلکس پروتئینی mRNA را در ناحیه پاییندست جایگاه پلی برش میدهد و mRNA از پلیمراز جدا میشود.
پردازش mRNA
در سلولهای پروکاریوتی mRNA بالافاصله پس از رونویسی و بدون تغییر به پروتئین ترجمه میشود. اما در سلولهای یوکاریوتی mRNA پس از تغییراتی از هسته خارج و ترجمه میشود. این تغییرات پایداری mRNA را افزایش میدهد. اضافه شدن $$5^prime CAP$$، اضافه شدن دم پلی آدنین و جدا شدن اینترونها سه تغییری است که پس از رونویسی روی mRNA ایجاد میشود.
- اضافه شدن $$5^prime CAP$$: همزمان با سنتز mRNA، کربن $$5^prime$$ نوکلئوتید ۷-متیل گوانوزین به فسفاتهای انتهای $$5^prime$$ متصل میشود. $$5^prime CAP$$ از تجزیه mRNA بهوسیله آنزیمهای نوکلئاز جلوگیری میکند.
- اضافه شدن دم پلی آدنین: پس از خاتمه رونویسی و جدا شدن mRNA آنزیم پلی A پلیمراز حدود ۲۰۰ نوکلئوتید آدنین به انتهای $$3^prime$$ اضافه میکند. این توالی علاوه بر جلوگیری از تجزیه RNA بهوسیله نوکلئازها، سیگنال خروج RNA از هسته است.
- جدا شدن اینترونها: قبل از ترجمه، اینترونها باید از mRNA جدا و اگزونها به هم متصل شود. این فرایند بهوسیله مجموعهای از پروتئینها و RNAهای کوچک هسته به نام اسپلاسوزوم انجام میشود. اولین نوکلئوتید انتهای $$5^prime CAP$$ هر اینترون GU و نوکلئوتیدهای انتهای $$3^prime$$ آن AG است. اسپلایسوزوم این نوکلئوتیدها را شناسایی میکند. در مرحله اول اسپلایسوزوم در انتهای $$$$ اینترون برش ایجاد میکند و این انتها به نوکلئوتید A نزدیک انتهای $$$$ متصل میشود. در مرحله دوم اسپلایسوزوم در انتهای $$$$ اینترون برش ایجاد و انتهای اگزونها را به هم متصل میکند.
ترجمه در ژنتیک چیست؟
در ترجمه کدونها به توالی آمینواسیدی در پروتئین ترجمه میشود. در مجموع ۶۴ کدون مختلف در mRNA وجود دارد که ۶۱ کدون به ۲۰ آمینواسید ترجمه میشود و ۳ کدون سیگنال پایان ترجمه است. AUG کدون متیونین و در تمام mRNAها کدون آغاز ترجمه است. به همین دلیل تمام زنجیرههای پلیپپتیدی اولیه با متیونین شروع میشود. به غیر از تریپتوفان و میتونین، بیش از یک کدون برای ترجمه آمینواسیدها وجود دارد. کدونهای یک آمینواسید در نوکلئوتید سوم با هم تفاوت دارند. برای مثال کدونهای سرین UCU و UCC و UCA و UCG است. ترجمه mRNA پروکاریوتها و یوکاریوتها در سه مرحله شروع، طویلسازی و خاتمه انجام میشود. اما فاکتورهای ترجمه و نوع ریبوزوم در این سلولها متفاوت است. در ادامه مطلب ژنتیک چیست مراحل ترجمه را با هم بررسی میکنیم.
شروع ترجمه
اولین مرحله ترجمه در پروکاریوتها و یوکاریوتها تشکیل کمپلکس شروع است. این کمپلکس در پروکاریوتها از زیرواحد ۳۰s ریبوزوم، mRNA الگو، سه فاکتور شروع IF1 و IF2 و IF3 و tRNA آغازگر (متیونین tRNA) تشکیل شده است. برای تشکیل کمپلکس شروع، IF3 مولکول mRNA را به زیرواحد ۳۰S ریبوزوم نزدیک و توالی شاین دالگارنو (AGGAGG) در بالادست کدون آغاز mRNA به rRNA ریبوزوم متصل میکند. در ادامه ریبوزوم برای پیدا کردن کدون آغاز روی mRNA حرکت میکند. IF2 متیونین فرمیل-tRNA را به زیرواحد ۳۰S ریبوزوم نزدیک میکند، tRNA در جایگاه P (پپتید | Peptide) ریبوزم قرار میگیرد و آنتیکدون آن با کدون AUG پیوند هیدروژنی تشکیل میدهد. IF2 یک GTPase است که با هیدرولیز GTP به GDP انرژی لازم برای اتصال زیرواحد بزرگ ریبوزوم (۵۰S) به زیرواحد کوچک را فراهم میکند. با اتصال زیرواحد بزرگ به زیرواحد کوچک فاکتورهای شروع ترجمه جدا میشود.
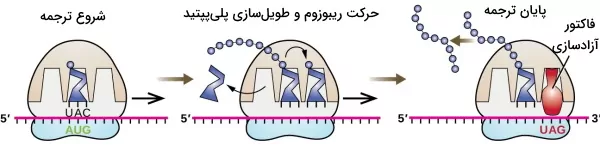
کمپلکس شروع ترجمه سلولهای یوکاریوتی از زیرواحد ۴۰S ریبوزوم، mRNA الگو، فاکتورهای ترجمه یوکاریوتی (eIFs)، CBPs (پروتئینهای اتصالی به CAP) تشکیل شده است. برای تشکیل این کمپلکس eIF3 به زیرواحد ۴۰S ریبوزوم متصل میشود. سپس eIF2 مولکول tRNA-متیونین را به ریبوزوم متصل میکند و در آخر eHFA-1 به این کمپلکس اضافه میشود. mRNA الگو بهوسیله eIF4 به ریبوزوم نزدیک و به آن متصل میشود. در یوکاریوتها والی شاین دالگارنو وجود ندارد و eIF4 و CBPs متیل گوانوزین انتهای $$5^prime$$ مولکول mRNA را شناسایی میکند. پس از تشکیل این کمپلکس، ریبوزوم برای پیدا کردن کدون شروع روی mRNA حرکت میکند.
پس از اتصال tRNA آغازگر به کدون شروع، فاکتورهای رونویسی از ریبوزوم جدا میشود. در این مرحله eIF6 زیرواحد بزرگ ریبوزوم را به زیروواحد کوچک تنزدیک میکند. eFI6 مهارکننده اتصال زیرواحد بزرگ به زیرواحد کوچک است. eFI5 پروتئین GTPase است که با هیدرولیز GTP به GDP، انرژی لازم برای جدا شدن eFI6 و اتصال دو زیرواحد ریبوزوم را فراهم میکند.
طویل سازی زنجیره پلیپپتیدی
طویلسازی، چرخهای از اتصال آمینواسید-tRNA، تشکیل پیوند پپتیدی و حرکت ریبوزوم است. در زیرواحد بزرگ ریبوزوم سه جایگاه برای tRNA وجود دارد. A جایگاه tRNA متصل به آمینواسید، P جایگاه tRNA متصل به زنجیره پلیپپتیدی و E جایگاه خروج tRNA است. در ابتدای طویلسازی tRNA آغازگر در جایگاه P قرار دارد و جایگاه A و E خالی از tRNA است. کدون دوم در جایگاه A قرار دارد. در پروکاریوتها آمینواسید بعدی بهوسیله EF-Tu و در یوکاریوتها بهوسیله eEF1-A به کدون متصل میشود. EF-Tu و eEF1 از GTPaseهای فرایند ترجمه است که با هیدرولیز GTP به GDP انرژی لازم برای اتصال tRNA به کدون را فراهم میکند. برای شروع دور بعدی EF-Ts مولکول GDP در EF-Tu را با GTP و eEF1-B مولکول GDP در eEF1-A را با GTP جایگزین میکند.
در مرحله دوم آنزیم پپتدیل ترانسفراز گروه کربوکسیل آمینواسید متصل به tRNA جایگاه P را جدا و به گروه آمین آمینواسید متصل به tRNA جایگاه A متصل میکند. این آنزیم از rRNA و پروتئینهای مختلف تشکیل شده است. پس از تشکیل پیوند پپتیدی، ریبوزوم به اندازه یک کدون جابهجا میشود. همزمان tRNA جایگاه P به جایگاه E و پپتیدیل-tRNA جایگاه A به جایگاه P منتقل میشود. در این مرحله EF-G (پروکاریوت) و eEF2 (یوکاریوت) با هیدرولیز GTP انرژی لازم برای حرکت ریبوزوم را فراهم میکند. در پایان این مرحله جایگاه E و A خالی و ریبوزوم آماده دور بعدی ترجمه است.
خاتمه ترجمه
در پایان ترجمه یکی از کدونهای UAG یا UAA یا UGA در جایگاه A قرار میگیرد. فاکتورهای رهاسازی (RFs) به جای آمینوآسیل-tRNA کدونهای پایان را شناسایی میکنند. در یوکاریوتها eRF1 تمام کدونهای پایان را شناسایی میکند. اما در پروکایوتها RF1 کدونهای UAA و UAG و RF2 کدونهای UAA و UGA را شناسایی میکند. در سلولهای یوکاریوتی eRF1 به کدون پایان و eRF2 به ریبوزم متصل میشود. در پروکاریوتها RF1 یا RF2 به کدون پایان و RF3 به ریبوزوم متصل میشود. فاکتورهای رهاسازی هیدرولیز پیوند زنجیره پلیپپتیدی با tRNA و جدا شدن دو زیرواحد ریبوزوم را کاتالیز میکنند.
جهش در ژنتیک چیست؟
به تغییرات ژنتیکی که منجر به تغییر عملکرد پروتئينها و ساختار پروتئین یا غیرفعال شدن آنها میشود، جهش گفته میشود. جهش ژنتیکی به دلیل تغییر یک یا دو نوکلئوتید یا تغییر تعداد و ساختار کروموزومها ایجاد شود. جهشهای نوکلئوتید به دلیل عوض شدن یک نوکلئوتید با نوکلئوتید دیگر، اضافه شدن یک نوکلئوتید به توالی ژن یا حذف یک نوکلئوتید از توالی ژن ایجاد میشود. اضافه و حذف شدن یک نوکلئوتید، چارچوب خوانش ژن را تغییر میدهد و توالی آمینواسیدهای پروتئین تغییر میکند. اما تغییر یک نوکلئوتید ممکن است یکی از کدونهای دیگر همان آمینواسید را ایجاد، کدون را به کدون آمینواسید دیگر تبدیل یا کدون آمینواسید را به کدون پایان تبدیل کند.
تعداد کروموزومها به دلیل صحیح جدا نشدن کروموزومهای همولوگ در آنافاز میتوز و میوز ایجاد میشود. جهشهای ساختاری کروموزوم به دلیل حذف قطعه از کروموزوم، جابهجایی قطعهای از DNA بین کروموزومهای غیرهمولوگ، شکسته و معکوس شدن قطعهای از DNA روی یک کروموزوم، دو یا چند برابر شدن یک ژن روی یک کروموزوم و اضافه شدن قطعهای از DNA به یک کروموزوم ایجاد میشود. اشتباهات همانندسازی، ترکیبات شیمیایی، پرتوهای یونیزاسیون، اشعه UV و گرما عوامل ایجاد جهشهای ژنتیکی هستند.
ترمیم جهش در ژنتیک چیست؟
در بخش قبلی مطلب ژنتیک چیست با انواع جهشهای ژنتیکی آشنا شدیم. در این بخش مکانیسمهای سلولی برای ترمیم این جهشها را مرور میکنیم. در سلولهای پروکاریوت و یوکاریوت مسیرهای آنزیمی برای ترمیم جهشهای DNA وجود دارد. ترمیم مستقیم، جدا کردن باز، جدا کردن نوکلئوتید، ترمیم ناجور باز، ترمیم شکست یک رشته DNA و ترمیم شکست دو رشته DNA مکانیسمهای سلول برای اصلاح جهشهای ژنتیکی است.
ترمیم مستقیم
در این روش DNA بدون جدا شدن یا جایگزینی نوکلئوتیدها ترمیم میشود. جهشهای ایجاد شده بهوسیله آلکیلاسیون بازها بهوسیله این روش ترمیم میشود. در این فرایند آنزیم $$O^6$$-آلکیل گوانین-DNA آلکیل ترانسفراز گروههای آلکیل را از بازها جدا میکنند و آنزیمهای داکسیژناز ساختار آلکیلها را تغییر میدهند. پروتئینهای شبه آلکیل ترانسفراز، با مهار این آنزیمها مسیر جدا کردن نوکلئوتید را برای ترمیم DNA آلکیله شده فعال میکنند.
جدا کردن باز
در این روش اکسیداسیون، دآمیناسیون، آلکیلاسیون و جایگاههای بدون باز DNA ترمیم میشود. این روش در مرحله G1 چرخه سلولی فعال است. در این روش آنزیمهای گلیکوزیلاز (آنزیمهای تجزیهکننده پیوند باز و قند) تغییر شکل ایجاد شده در ساختار DNA به دلیل جهش را شناسایی میکنند. این آنزیمها باز را از نوکلئوتید جدا میکنند. پس از جدا شدن باز بهوسیله این آنزیمها، آنزیم اندونوکلئاز پیوند فسفودیاستری انتهای $$5^prime$$ نوکلئوتید بدون باز را برش میدهد و قند باقیمانده از رشته DNA جدا میشود. سپس آنزیم DNA پلیمراز بتا جایگاه بدون نوکلئوتید را پر میکند و آنزیم لیگاز بین نوکلئوتید قبلی و نوکلئوتید جدید پیوند ایجاد میکند.
جدا کردن نوکلئوتید
این روش برای ترمیم جهشهای ایجاد شده بهوسیله ترکیبات شیمیایی و اشعه UV استفاده میشود. در این روش آنزیم هلیکاز دو رشته DNA را از هم باز میکند و آنزیمهای اندونوکلئاز پیوند فسفودیاستری نوکلئوتیدها را در فاصله چند بازی از نوکلئوتید نغییریافته برش میدهد. در نتیجه قطعهای از DNA جدا میشود و آنزیم پلیمراز با الگو قرار دادن رشته مقابل فاصله بین نوکلئوتیدها را پر میکند.
ترمیم ناجور باز
این روش ترمیم بلافاصله پس از پایان همانندسازی بازهای مکمل اشتباه، اضافه شدن یا حذف شدن باز را ترمیم میکند. در مرحله اول این روش پروتئینهای ترمیمی ناجور باز را شناسایی میکنند. سپس آنزیم اگزونوکلئاز در فاصله چند بازی جایگاه جهش برش ایجاد و نوکلئوتیدها را خارج میکند. در آخر آنزیم پلیمراز با الگو قرار دادن رشته سالم، فاصله بین نوکلئوتیدها را پر میکند.
ترمیم شکست یک رشته DNA
اکسیداسیون، نوکلئوتید بدون باز و خطاهای آنزیم توپوایزومراز در همانندسازی منجر به ایجاد DNA تکرشتهای میشود. اولین مرحله این روش اتصال پروتئینهای PARP1 به DNA تکرشتهای برای جلوگیری از تجزیه آن بهوسیله آنزیمهای نوکلئازی است. د رماحل بعدی این شکست بهوسیله ترمیم ناجورباز، جدا کردن نوکلئوتید یا جدا کردن باز ترمیم میشود.
ترمیم شکست دو رشته DNA
شکستهای دو رشته DNA بیشتر به دلیل تابش پرتوهای یونیزاسیون ایجاد میشود. تابشهای یونیزاسیون در بیشتر موارد انتهای تکرشتهای در محل شکست ایجاد میکنند. سلول برای ترمیم این شکستها از مکانیسم اتصال انتهای غیر همولوگ و نوترکیبی همولوگ استفاده میکند.
- اتصال انتهای غیرهمولوگ: این روش در تمام مراحل چرخه سلولی برای ترمیم DNA استفاده میشود اما در G1 اهمیت بیشتری دارد. در این روش بخشی از اطلاعات ژنتیکی در DNA ترمیم شده وجود ندارد. در مرحله اول مجموعهای از پروتئینهای نوترکیبی میوز دو انتهای DNA در محل شکست را شناسایی میکنند و و آنزیمهای سرین-تروئونین کیناز به این پروتئینها متصل میشوند. آنزیمهای کیناز به سرین هیستونها گروه فسفات اضافه میکند. اضافه شدن گروه فسفات کنفورماسیون هیستون را تغییر میدهد و هیستون به پروتئینهای دیگر متصل میشود. این پروتئینها فاکتور رونویسی p53 و p53 رونویسی ژن پروتئینها و آنزیمهای ترمیم DNA (پروتئینهای Ku، آنزیمهای DNA کیناز، نوکلئازها و لیگاز) را فعال میکند. آنزیمهای نوکلئاز دو انتهای تکرشتهای محل شکست DNA را برش میدهند و دو انتهای دورشتهای ایجاد میشود. در آخر آنزیم لیگاز انتهای دو رشته را به هم متصل میکند.
- نوترکیبی همولوگ: در این روش از کروماتیدهای خواهری به عنوان رشته الگو برای ترمیم رشته آسیبدیده استفاده میشود. به همین دلیل در مرحله G1 که کروماتیدهای خواهری وجود ندارد، سلول نمیتواند از این روش استفاده کند. این روش دقیقتر از اتصال انتهای غیرهمولوگ است و تمام اطلاعات ژنتیکی در DNA ترمیم شده وجود دارد. توالی نوکلئوتیدها در رشته DNA ایجاد شده در این روش ممکن است کاملا شبیه DNA اولیه نباشد اما تغییری در عملکرد آن ایجاد نمیشود. در مرحله اول این روش ترمیم شکست DNA بهوسیله پروتئینهای مشابه اتصال انتهای غیرهمولوگ شناسایی میشود. این پروتئینها به هیستون گروههای فسفات اضافه میکنند، p53 فعال میشود و ژنهای آنزیمهای ترمیمی فعال میشود. در مرحله بعد آنزیمهای نوکلئاز در فاصله ۱۰۰ تا ۲۰۰ جفتبازی محل شکست در رشتهای که انتهای $$5^prime$$ آن در محل شکست قرار دارد، برش ایجاد و نوکلئوتیدها را جدا میکند. در نتیجه انتهای تکرشتهای بلندی در محل شکست ایجاد میشود. در این مرحله پروتئینهای RPA به DNA تکرشتهای متصل میشود تا از تجزیه آنزیمی این رشته جلوگیری کند. در مرحله آخر آنزیم پلیمراز کروماتید خواهری یا کروموزوم غیرهمولوگ را الگو قرار میدهد و رشتهای که انتهای $$3^prime$$ آن در محل شکست قرار دارد، همانندسازی میشود. در نهایت آنزیم لیگاز انتهای $$3^prime$$ و $$5^prime$$ را به هم متصل میکند.

نوترکیبی در ژنتیک چیست؟
در نوترکیبی ژنتیکی دو قطعه نوکلئوتیدی در یک مولکول DNA یا بین دو مولکول DNA جابهجا میشود. نوترکیبی بیشتر بین توالیهای همولوگ کروموزومها و در پروکاریوتها و یوکاریوتها ایجاد میشود. نوترکیبی علاوه بر ایجاد تنوع ژنتیکی در ترمیم DNA آسیبدیده نقش دارد. در قارچها، گیاهان و جانوران نوترکیبی در تقسیم میوز ایجاد میشود. سلولهای باکتری بهوسیله تقسیم دوتایی تکثیر میشود. در این تقسیم نوترکیبی وجود ندارد. به همین دلیل برخلاف موجوداتی که با تقسیم میوز تکثیر میشوند، نوترکیبی باکتری از والدین به نسل بعدی منتقل نمیشود و بین افراد یک نسل ایجاد میشود. نوترکیبی در پروکاریوتها بهوسیله در پروکاریوتها در ترانسداکشن و کانجوگاسیون ایجاد میشود. نوترکیبی در چهار مرحله کنار هم قرار گرفتن دو کروموزوم همولوگ، شکسته شدن بخشی از مولکول DNA، تبادل ژنی بین رشتهها و اتصال قطعه نوترکیب به رشته قبلی انجام میشود.
نوترکیبی میوز
در تقسیم میوز سلولهای زاینده دیپلوئید به گامتهای هاپلوئید تقسیم میشود. در پروفاز I میوز کروموزومهای همولوگ بهوسیله پروتئینهای کمپلکس سیناپتونمال کنار هم قرار میگیرد. در این حالت بخشی از یک کروموزوم ممکن است روی کروموزوم دیگر قرار بگیرد و بین آنها همپوشانی ایجاد شود. به این اتفاق کراسینگاور کروموزومها گفته میشود. کروموزومها در محل کراسینگاور شکسته و قطعات DNA بین آنها مبادله میشود. در نتیجه آرایش ژنها در گامتها با سلولهای زاینده متفاوت است.
نوترکیبی در ترانسداکشن
نوترکیبی در ترانسداکشن به کمک باکتریوفاژ ایجاد میشود. باکتریوفاژها ویروسهایی هستند که از یک پوشش پروتئینی و مولکول DNA تشکیل شدهاند. این ویروسها برای آلوده کردن باکتری DNA خود را از غشای پلاسمایی به سیتوپلاسم باکتری تزریق میکنند. از آنجایی که ویروسها آنزیمهای لازم برای همانندسازی، رونویسی و ترجمه را ندارند، DNA آنها وارد بخشی از DNA باکتری میشود. پس از چند بار تکثیر و سنتز پروتئینها DNA ویروسی از DNA باکتری جدا میشود و در پوشش پروتئینی قرار میگیرد. بخشی از DNA باکتری ممکن است همراه DNA ویروسی جدا شود. ویروس کامل با لیز شدن باکتری آزاد میشود. ویروس باکتریهای جدید را آلوده میکند و بخشی از DNA باکتری قبلی به باکتری جدید منتقل میشود.
نوترکیبی در کانجوگاسیون
در کانجوگاسیون DNA که معمولا پلاسمید است، مستقیم از یک باکتری به باکتری دیگر منتقل میشود. برای کانجوگاسیون دو سلول باکتری به هم نزدیک و بهوسیله پیلوس جنسی به هم متصل میشوند. پیلوس رشتهای پروتئینی در سطح باکتری است که پروتئینهای آن در ژن F (ژن زایش | Fertility) باکتری اهداکننده کد میشود.
نوترکیبی مکان ویژه
نوترکیبی مکان ویژه در یک نقطه مشخص دو DNA ایجاد میشود. برخلاف سایر مکانیسمهای نوترکیبی، جهت قرار گرفتن توالیها مهم است. این توالی از ۳۰ تا ۲۰۰ نوکلئوتید تشکیل شده است و معمولا دو موتیف با تکرارهای معکوس در آن وجود دارد که بهوسیله آنزیم ریکامبیناز شناسایی میشود. ادغام DNA باکتریوفاژها با DNA باکتری یکی از مثالهای نوترکیبی مکان ویژه است. «آنزیمهای ریکامبیناز مکان ویژه» (Site Specific Recombinase | SSR) توالیهای مشخص DNA را شناسایی میکنند و به DNA متصل میشوند. این آنزیمها به دو دسته سرین ریکامبینازها و تیروزین ریکامبینازها تقسیم میشوند.در جایگاه فعال سرین کامبینازها، آمینواسید سرین و در جایگاه فعال تیروزین ریکامبینازها، آمینواسید تیروزین قرار دارد. مکانیسم نوترکیبی این دو آنزیم با هم متفاوت است.
- سرین ریکامبیناز: گروه OH سرین جایگاه فعال این آنزیمها به پیوند فسفودیاستری بین نوکلئوتیدها در دو مولکول DNA حمله و در دو رشته شکست ایجاد میکند. در مرحله بعد رشتههای دو DNA جابهجا میشود و در مرحله آخر آنزیم ریکامبیناز پیوند قطعه جدید و قطعه قدیمی پیوند ایجاد میکند.
- تیروزین ریکامبیناز: گروه OH تیروزین ابن آنزیمها به گروه فسفات پیوند فسفودیاستر یکی از رشتهها در دو مولکول DNA حمله میکند. رشتههای شکسته شده در دو مولکول با هم جابهجا میشوند. در مرحله دوم رشته دیگر شکسته و جابهجا میشود. در پایان آنزیم ریکامبیناز بین نوکلئوتیدهای قطعه جدید و قدیمی پیوند ایجاد میکند.
اصول ژنتیک مندلی
اگر در بخشهای قبلی مطلب ژنتیک چیست با ما همراهی کرده باشید، متوجه شدید که ژنها قطعاتی از DNA هستند که اطلاعات ژنتیکی تولید یک پروتئین خاص را ذخیره میکنند و از فشرده شدن DNA کروموزوم ایجاد میشود. در این بخش نحوه به ارث رسیدن ژنها را بررسی میکنیم. «گریگور مندل» (Johann Gregor Mendel) اولین فردی بود که در قرن ۱۹ میلادی نحوه به ارث رسیدن ژنها را بررسی کرد. نتیجه تحقیقات این دانشمند پایه ژنتیک کلاسیک یا ژنتیک مندلی است. مندل با بررسی حدود ۳۰،۰۰۰ گیاه نخودفرنگی به سه قانون غالبیت، قانون تفکیک ژنها و جور شدن مستقل ژنها رسید که نحوه به ارث رسیدن و بروز صفات را نشان میدهد.
- قانون غالبیت: بر اساس قانون غالبیت در موجودات دیپلوئید اللها با هم رابطه غالب و مغلوبی دارند. در موجودات دیپلوئید هر الل از یکی از والدین به ارث میرسد. در این حالت اگر فرض کنیم A و B دو الل یک ژن رنگ نخودفرنگی است که بیان الل A سبب سبز شدن رنگ نخودفرنگی و بیان الل B سبب زرد شدن رنگ نخود فرنگی میشود. به علاوه الل A بر الل B غالب است. اگر حتی یک الل A در سلولهای نخودفرنگی باشد، رنگ آن سبز میشود.
- قانون تفکیک ژن: بر اساس این قانون اللهای یک ژن در تقسیم میوز از هم جدا میشوند و در پایان میوز هر الل در یکی از گامتها وجود دارد. در نتیجه از هر والد یک الل به فرزند منتقل میشود.
- قانون جور شدن مستقل ژنها: بر اساس این قانون اللهای ژنهای مختلف در گامتزایی به طور مستقل از هم جدا میشوند.
بسط قانون غالبیت
تمام ژنها بر اساس قوانین مندل به ارث نمیرسد و بروز صفات در تمام ژنها از قوانین مندل پیروی نمیکند. بر این اساس ارتباط بین اللهای یک ژن علاوه بر غالب و مغلوبی ممکن است بارزیت ناقص یا همتوانی باشد. در رابطه بارزیت ناقص، فرزندان هتروزیگوت صفتی را نشان میدهند که ترکیبی از صفات والدین است. برای مثال یکی رنگ گلبرگهای والد نر سفید و رنگ گلبرگهای والد ماده قرمز است. اما از آمیزش آنها گلهای با گلبرگهای قرمز، سفید یا صورتی به وجود میآید. در رابطه همتوانی، بعضی از فرزندان صفت هر دو والد را نشان میدهند. برای مثال اگر گروه خونی یکی از والد انسان A و دیگری B باشد، فرزندان هتروزیگوت گروه خونی AB دارند.

به علاوه بعضی از ژنها بیش از دو الل دارند. رابطه بین این اللها ممکن است غالب و مغلوبی، بارزیت ناقص یا همتوانی باشد. برای مثال ژن گروه خونی انسان سه الل A و B و O دارد که در ژنوم هر فرد دو الل آن وجود دارد و رابطه آنها همتوانی است. بروز بعضی از صفات به چند ژن وابسته است. برای مثال قد، رنگ پوست و رنگ مو صفاتی چند ژنی انسان هستند. همچنین شرایط محیطی نحوه بیان ژنها را تغییر میدهد. برای مثال فنوتیپ رنگ موی هیمالیایی در خرگوشها، صفت وابسته به دمای محیط است. ژن رنگدانه موی خرگوش در دمای محیطی کم و زیاد بیان میشود. اما ساختار پروتئین این ژن در دماهای بالا به درستی شکل نمیگیرد و رنگ موها تغییر میکند.
نقض قانون تفکیک ژن ها
ژنهای وابسته به کروموزوم x مثال نقض قانون تفکیک ژنهای مندل است. در انسان، بسیاری از جانوران و بعضی از گیاهان جنسیت بهوسیله گروموزومهای X و Y مشخص میشود. در بخشهای قبلی این مطلب از مجله فرادرس توضیح دادیم که ژنوتیپ فرد ماده XX و ژنوتیپ فرد نر XY است. بیماری هموفیلی یکی از بیماریهای ژنتیکی مغلوب است که ژن آن روی کروموزوم X قرار دارد. در نتیجه افراد نر تنها یک الل هموفیلی از یکی از والدین دریافت میکنند و اگر این الل مغلوب باشد به بیماری مبتلا میشوند. اما افراد ماده یک الل از هر والد دریافت میکنند که ممکن است غالب یا مغلوب باشد و تنها افرادی که هموزیگوت مغلوب هستند به بیماری مبتلا میشوند.
نقض قانون جور شدن مستقل ژن ها
قانون جور شدن مستقل ژنها در مورد ژنهایی درست است که روی کروموزومهای متفاوتی قرار دارند. اما ژنهایی که روی یک کروموزوم قرار دارند به هم پیوسته هستند و با هم به ارث میرسد. این ژنها تنها در صورتی که با نوترکیبی از هم جدا شوند از قانون جور شدن مستقل ژنها پیروی میکنند.
مهندسی ژنتیک چیست؟
مهندسی ژنتیک مجموعهای از تکنیکهای آزمایشگاهی است که برای ایجاد تغییر در ساختار DNA و ایجاد فنوتیپهای جدید استفاده میشود. در این روشها یک ژن یا تعدادی ژن جدید به ژنوم یک سلول یا موجود زنده منتقل میشود. ژن مورد نطر ممکن است از افراد یک گونه انتخاب شود و پس از تغییرات به افراد همان گونه منتقل شود یا از افراد یک گونه به گونه دیگر منتقل شود. از این روش میتوان برای تولید واکسن نوترکیب، اصلاح نژاد گیاهان و محصولات کشاورزی، تولید داروهای پروتئینی و ژندرمانی استفاده کرد. برای مثال با این روش میتوان گیاهانی تولید کرد که مقاومت بیشتری در تنشهای آبی دارند. استخراج DNA، جداسازی DNA، تکثیر DNA و کلونینگ مراحل مهندسی ژنتیک است.
استخراج DNA در مهندسی ژنتیک چیست؟
برای ایجاد تغییر در DNA و جدا کردن ژن مورد نظر اولین مرحله خارج کردن DNA از سلول است. در تمام تکنیکهای استخراج DNA غشای پلاسمایی لیز و ماکرومولکولهای دیگه بهوسیله آنزیم تجزیه میشود. برای لیز کردن غشای پلاسمایی از دترجنتهای بافری استفاده میشود. حذف آنزیمی مولکولهای دیگر ازجمله RNA و پروتئینها از آلودگی نمونه DNA و تجزیه آن بهوسیله نوکلئازها جلوگیری میکند. در مرحله بعد DNA بهوسیله الکل و سانتریفوز از سایر ترکیبات سلولی جدا میشود. مولکول DNA در این مرحله یک توده ژلهای است. RNA سلول را هم میتوان با این روش استخراج کرد. با این تفاوت که RNA بسیار ناپایدارتر ار DNA است و آنزیمهای تجزیه آن در طبیعت زیاد است. حتی پوست دست ما آنزیمهای RNAase ترشح میکند.
جداسازی DNA در مهندسی ژنتیک چیست؟
DNA مولکولی است که در pH خنثی و قلیایی بار منفی دارد. به همین دلیل در میدان الکترکی حرکت میکند. ژل الکتروفورز تکنیکی است که برای جدا کردن قطعات DNA بر اساس بار و اندازه از هم جدا میکند. به کمک این تکنیک میتوان یک کروموزوم کامل یا قطعات DNA را از هم جدا کرد. در ژل الکتروفوز نمونه DNA در چاهکهایی نزدیک قطب منفی میدان الکتریکی ریخته میشود. پس از ایجاد جریان الکتریکی مولکولها از قطب منفی به قطب مثبت حرکت میکنند. مولکولهای ککوچکتر زودتر از منافذ ژل عبور میکنند و به انتهای مثبت ژل نزدیکتر هستند. اما مولکولهای بزرگتر کندتر حرکت میکنند و به انتهای منفی ژل نزدیکتر هستند. برای دیدن جایگاه DNA روی ژل از تکنیکهای مختلف ازجمله رنگآمیزی استفاده میشود.
تکثیر DNA در مهندسی ژنتیک چیست؟
مقدار DNA استخراج شده معمولا برای انجام تکنیکهای مختلف کافی نیست. در این حالت از واکنش زنجیره پلیمراز یا PCR برای تکثیر تمام یا قطعهای از DNA استفاده کرد. در این روش از آنزیم مقاوم به دما به نام Taqپلیمراز، پرایمرهای RNA و نوکلئوتیدها استفاده میشود. هر دور RCR از در سه مرحله باز شدن دو رشته DNA از هم، اتصال پرایمر و همانندسازی DNA انجام میشود. از این روش علاوه بر تکثیر DNA میتوان برای تشخیص بیماریها، مقایسه DNA دو گونه و مشخص کردن توالی نوکلئوتیدی در یک قطعه از DNA استفاده کرد.
کلونینگ در مهندسی ژنتیک چیست؟
کلونینگ یا شبیهسازی در زیستشناسی، تولید یک کپی ژنتیکی از یک سلول یا تمام بدن یک موجود زنده است. از این روش میتوان برای تکثیر DNA یا برری بیان ژن استفاده کرد. در این روش DNA استخراج شده بهوسیله یک ناقل یا مستقیم به یک سلول باکتری، قارچ یا پستاندار منتقل میشود. اولین مرحله کلونینگ انتخاب و جدا کردن ژن مورد نظر از DNA است. در مرحله بعد ژن مورد نظر به حامل یا وکتور منتقل میشود. در ادامه وکتور به سلولهای هدف منتقل میشود و در مرحله آخر محصول کلونینگ (DNA یا پروتئین) استخراج میشود. برای جدا کردن ژن میتوان از PCR یا آنزیمهای محدوکننده استفاده کرد. در روش PCR میتوان از پرایمرهایی استفاده کرد که دو طرف ژن مورد نظر قرار میگیرند. در نتیجه تنها قطعهای از DNA تکثیر میشود. آنزیمهای محدودکننده اندونوکلئازهای باکتریایی هستند که توالی مشخصی از DNA را برش میدهند.
اگر میزبان باکتری باشد از پلاسمیدها میتوان به عنوان حامل ژن استفاده کرد. پلاسمیدها DNA حلقوی باکتری هستند که مستقل از کروموزوم اصلی تکثیر میشود. برای اتصال زن مورد نظر به پلاسمید، هر دو DNA با یک آنزیم محدودکننده برش داده میشود. سپس دو DNA با هم مخلوط و انتهای آنها بهوسیله آنزیم لیگاز به هم متصل میشود. پلاسمید را میتوان با استفاده از ایجاد تنشهای فیزیکی، «تزریق میکرو» (Microinjection)، «الکتروپورشن» (Electroporation)، «سونیکاسیون» (Sonication)، تفنگ ژنی، باکتریوفاژ یا لیپوزوم به سلول مورد نظر منتقل کرد. در نهایت DNA مورد نظر بهوسیله سیستم مولکولی میزبان تکثیر یا بیان میشود.
ژنتیک جمعیت چیست؟
ژنتیک جمعیت بخشی از ژنتیک است که پراکندگی و تغییر فراوانی ژنوتیپ و فنوتیپ را در یک جمعیت یا بین جمعیتهای مختلف بررسی میکند. انتخاب طبیعی، رانش ژنی، جهش و شارش ژنی تغییر میکند.
- انتخاب طبیعی: انتخاب طبیعی فرایندی است که به افزایش سازگاری موجودات زنده با محیط اطراف کمک میکند. در این فرایند ژنوتیپهایی که سازگاری فرد با محیط و شانس بقای آن را افزایش میدهند از نسلی به نسل دیگر منتقل میشود. برای مثال فرض کنید موشهایی با رنگ بدن مشکی و سفید وارد محیط جدیدی شدهاند که رنگ صخرههای آن مشکی است. در این حالت موشهای مشکی شانس بیشتری برای فرار کردن از شکارچی به دلیل همرنگ بودن با محیط دارند. در نتیجه میتوان گفت رنگ موش مشکی در این محیط یک صفت برتر است که شانس بقای جانور را افزایش میدهد. این صفت به نسلهای بعدی منتقل میشود و فراروانی فنوتیپ جمعیت پس از چند نسل تغییر میکند.
- رانش ژن: رانش ژنی تغییرات ایجاد شده در زنوتیپ و فنوتیپ جمعیت به دلیل اتفاقات تصادفی ازجمله بلایای طبیعی یا مهاجرت موجودات ایجاد میشود. این پدیده در تمام جمعیتهای محدود ایجاد میشود اما احتمال آن در جمعیتهای کوچکتر بیشتر است. در این نوع تغییرات برخلاف انتخاب طبیعی الل باقیمانده ممکن است منجر به افزایش یا کاهش بقای موجودات زنده شود. اثر گردن بطری یا گذرگاه باریک و اثر بنیانگذار دو مکانیسم رانش ژنی است.
- اثر گردن بطری: اثر گردن بطری در جمعیتهای بسیار کوچک ایجاد میشود. بالایای طبیعی (سیل، زلزله، آتشسوزی یا رانش زمین) پدیدههایی هستند که به شکل تصادفی تعداد زیادی از افراد یک جمعیت را از بین میبرد. در این شرایط فراوانی اللها در جمعیت بسیار تغییر میکند و حتی ممکن است بعضی از اللها حذف شود. برای درک بهتر این اثر فرض کنید تعداد زیادی تیله در یک بطری ریخته شده که دهانه باریکی دارد. هر بار که بخواهید تعدادی از تیلهها را از بطری به یک ظرف دیگر بریزید، تعداد کمی از تیلهها به شکل تصادفی داخل ظرف ریخته میشود و ترکیب رنگ تیلههای داخل ظرف با تیلههای بطری کاملا متفاوت است.
- اثر بنیانگذار: اثر بنیانگذار زمانی ایجاد میشود که گروهی از افراد یک جمعیت جدا میشوند و کلونی جدیدی تشکیل میدهند. در این حالت فراوانی اللها در کلونی تشکیل شده با جمعیت اولیه متفاوت است و حتی ممکن است بعضی از اللهای جمعیت اولیه در کلونی جدید وجود نداشته باشد.
- جهش: جهشهای ژنتیکی با تغییر یک یا چند نوکلئوتید یک ژن و تغییر ساختار یا تعداد کروموزومها منجر به ایجاد تغییرات ژنوتیپ و فنوتیپ ردر یک جمعیت میشود.
- شارش ژنی: شارش ژنی انتقال اللهای ژنتیکی بین دو جمعیت از یک گونه است که به دلیل مهاجرت افراد بین جمعیتها ایجاد میشود. این پدیده تنوع ژنتیکی در هر جمعیت را افزایش و تفاوتهای ژنتیکی بین دو جمعیت را کاهش میدهد.

اپی ژنتیک چیست؟
در بخشهای قبلی مطلب ژنتیک چیست توضیح دادیم که در تمام سلولهای بدن انسان ۴۶ کروموزوم یکسان وجود دارد که شکل ظاهری و مسیرهای بیوشیمیایی سلول را تعیین میکند. پس چرا انسولین فقط از سلولهای پانکراس و آنتیبادیها از پلاسموسیتها ترشح میشود. در مسیرهای تنظیمی سلول، ساختار DNA بدون تغییر توالی نوکلئوتیدها تغییر میکند. اپیژنتیک بخشی از ژنتیک است که این تغییرات را بررسی میکند. این تغییرات منجر به خاموش شدن ژنها و تمایز سلولهای مختلف میشود. متیلاسیون DNA، تغییرات هیستون و خاموش شدن ژنهای وابسته به RNAهای غیرکدکننده سه تغییر اپیژنتیکی سلولها است.
- متیلاسیون DNA: در متیلاسیون DNA گروههای متیل بهوسیله آنزیم متیل ترانسفراز به نوکلئوتید سیتوزین در توالی سیتوزین-گوانین (CpG) اضافه میشود که با توالیهای CpG اطرافش جزیره CG تشکیل میدهد. این توالی در پروموتر بیشتر ژنها وجود دارد. اضافه شدن گروه متیل به سیتوزین سبب اتصال فاکتورهای مهارکننده بیان ژن به DNA و مهار ژن میشود. به علاوه متیلاسیون DNA منجر به اتصال محکمتر هیستون به پروتئین و تشکیل یوکروماتین میشود. در بسیاری از سرطانها پروموتر ژنهای مهارکننده تومور متیله شده است.
- تغییرات هیستون: اضافه شدن گروههای متیل، استیل، فسفات و یوبیکوئیتین به هیستونها برهمکنش این پروتئینها با DNA را تغییر میدهد. گروه استیل معمولا به آمینواسیدهای لیزین هیستون متصل میشود و با کاهش بار مثبت هیستون برهمکنش هیستون-DNA را کاهش میدهد. در نتیجه رونویسی ژنها افزایش مییابد. اضافه شدن گروه متیل به هیستون بار این پروتئین را تغییر نمیدهد. اما بر اساس محل متیلاسیون رونویسی ژن را افزایش یا کاهش میدهد. یک تا سه گروه متیل به آمینواسید لیزین و یک تا ددو گروه به آمینواسید آرژینین این پروتئینها اضافه میشود. برای مثال متیلاسیون لیزین چهار هیستون H3 رونویسی را فعال و متیلاسیون لیزین ۲۷ هیستون H3 رونویسی را مهار میکند. گروههای فسفات به دم کربوکسیل هیستونها اضافه میشود. اضافه شدن H2A در فعال شدن سیستم ترمیم DNA نقش دارد. یوبیکوئیتین به لیزین هیستونها اضافه میشود و در خاموش شدن ژن نقش دارد.
- خاموش شدن ژنهای وابسته به RNAهای غیرکدکننده: RNAهای غیرکدکننده، مولکولهایی هستند که رونویسی میشوند اما ترجمه نمیشوند. miRNA (میکرو RNA) و siRNA (مولکول RNA تداخلی کوچک) انواع RNAهای غیرکدکننده و کوتاهتر از ۳۰ نوکلئوتید هستند. RNAهای غیرکننده بلند از ۲۰۰ یا تعداد بیشتری نوکلئوتید تشکیل میشود. این مولکولها تشکیل یوکروماتین و خاموش شدن ژن را تحریک میکنند.
سیتوژنتیک چیست؟
سیتوژنتیک آخرین گرایشی است که در مطلب ژنتیک چیست توضیح میدهیم. سیتوژنتیک بخشی از علم ژنتیک است که ساختار و ویژگیهای کروموزوم و رفتار و تغییرات آن در تقسیم میتوز و میوز را به همراه تاثیر آن بر فنوتیپ افراد بررسی میکند. تعیین کاریوتایپ، «هیبریداسیون فلورسنت در جا» (Fluorescent in situ hybridisation | FISH) و هیبریداسیون مقایسهای ژنوم سه تکنیک اصلی در سیتوژنتیک است.
- تعیین کاریوتایپ: در این روش کروموزومهای متافازی از سلول جدا و با روشهای استاندارد رنگآمیزی میشود. یوکروماتین و هتروکروماتینهای کروموزوم برهمکنشهای متفاوتی با رنگ دارند. به همین دلیل پس از رنگآمیزی در کروموزومها خطوط تیره و روشن دیده میشود. پس از رنگآمیزی از کروموزومها زیر میکروسکوپ عکس گرفته میشود. پس از رنگآمیزی کروموزومها از بزرگ به کوچک و به شکلی که سانترومر آنها روی یک خط افقی فرضی، بازوی کوتاه بالای خط و بازی کوتاه پایین خط باشد، مرتب میشوند.
- FISH: در این تکنیک از قطعات DNA نشانهگذاری شده با رنگهای فلوسنت (پروب) استفاده میشود که مکمل توالیهای DNA هستند. به کمک این روش میتوان حذف یا اضافه شدن قطعهای DNA در کروموزوم را شناسایی کرد. در این روش DNA پس از دناتوره شدن با گرما یا ترکیبات شیمیایی، با پروب مخلوط میشود. پروب با توالی DNA مکمل خود جفت میشود و میتوان آن را زیر میکروسکوپ فلورسنت مشاهده کرد.
- هیبریداسیون مقایسهای ژنوم: از این تکنیک برای مضاعفشدگی ژنها استفاده میشود. در این حالت از یک ژن دو یا چند نسخه روی یک کروموزوم وجود دارد. در این روش DNA نمونه و یک DNA کنترل وجود دارد که بهوسیله رنگهای فلورسنت متفاوت نشانهگذاری شده است. دو مولکول DNA پس از دناتوراسیون با هم مخلوط میشوند. شدت نور فلورسنت نمونه نسبت به کنترل برای هر کروموزوم رسم میشود و تعداد نسخههای ژنی را مشخص میکند. یکی از محدودیتهای CGH این است که تغییرات ژنتیکی را فقط در قطعات حدود ۵ تا ۱۰ مگابازی شناسایی میکند.
جمعبندی
در این مطلب از مجله فرادرس توضیح دادیم که ژنتیک بخشی از زیستشناسی است که چگونگی به ارث رسیدن صفات بین موجودات مختلف را توضیح میدهد. اطلاعات ژنتیکی موجودات در ماکرومولولهای DNA ذخیره و به نسل بعضی منتقل میشود. این مولکولها از دو رشته با زیرواحدهای نوکلئوتیدی تشکیل شدهاند و پس از فشرده شدن به آنها کروموزوم گفته میشود. در تقسیم سلولی از هر مولکول DNA دو مولکول جدید سنتز میشود که یک رشته DNA اولیه و یک رشته جدید دارند. آسیب DNA منجر به تغییر توالیهای نوکلئوتیدی از بین رفتن بخشی از اطلاعات ژنتیکی میشود. به این اتفاق جهش ژنتیکی گفته میشود. جهشها ممکن است تغییری در ویژگیهای موجود زنده ایجاد نکنند یا با ایجاد صفات جدید همراه شود. این صفات ممکن است یک بیماری باشد. سلول از مکانیسمهای ترمیم مستقیم، ترمیم ناجورباز، جدا کردن باز، جدا کردن نوکلئوتید، اتصال دو انتهای غیرهمولوگ و نوترکیبی همولوگ برای ترمیم جهشها استفاده میکند.
در ادامه مطلب ژنتیک چیست توضیح دادیم که در فرایند رونویسی از DNA یک نسخه RNA تهیه میشود. RNA با کمک ریبوزومها به پروتئین ترجمه میشود. اطلاعات پروتئینها در واحدهایی به نام ژن ذخیره شده است. به شکلهای مختلف یک ژن الل گفته میشود. ارتباط بین اللهای یک ژن در بروز صفات نقش دارد. ارتباط بین جهشها ممکن است غالب و مغولب، همتوانی و بارزیت ناقش باشد. در رابطه غالب و مغلوبی وجود یک الل ایجاد صفا الل دیگر را مهار میکند. در رابطه همتوانی صفت ایجاد شده صفت هر دو الل همزمان و به طور یکسان بروز میکند. اما در رابطه بارزیت ناقص صفت ایجاد شده ترکیبی از صفت دو الل است.
در ادامه توضیح دادیم که روشهای نوترکیبی علاوه بر ترمیم ژنها در ایجاد تنوع ژنتیکی نقش دارند. در بخشهای بعدی اصول ژنتیک مندلی و قوانین مندلی که در مطالعات بعدی نقض شد را مرور کردیم. در انتهای مطلب ژنتیک چیست ژنتیک جمعیت، سیتوژنتیک، اپیژنتیک و روشهای مهندسی ژنتیک را بررسی کردیم.
source